

Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage
- PMID: 29160308
- PMCID: PMC5726555
- DOI: 10.1038/nature24644
Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage
Erratum in
-
Publisher Correction: Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage.Nature. 2018 Jul;559(7714):E8. doi: 10.1038/s41586-018-0070-x. Nature. 2018. PMID: 29720650
Abstract
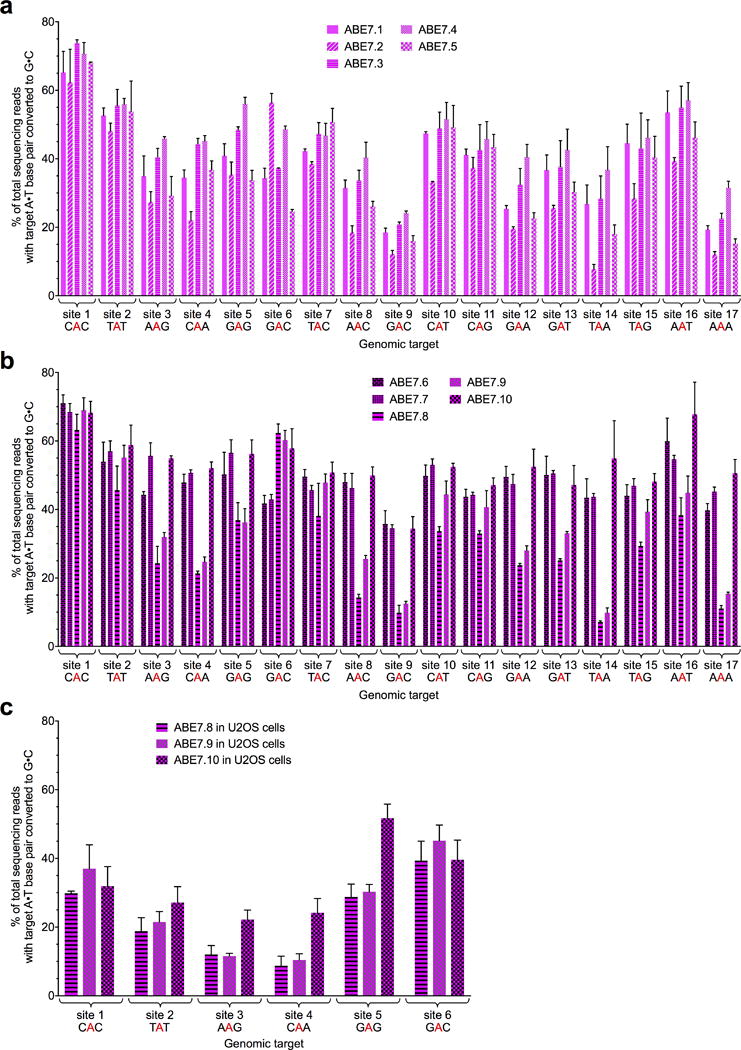
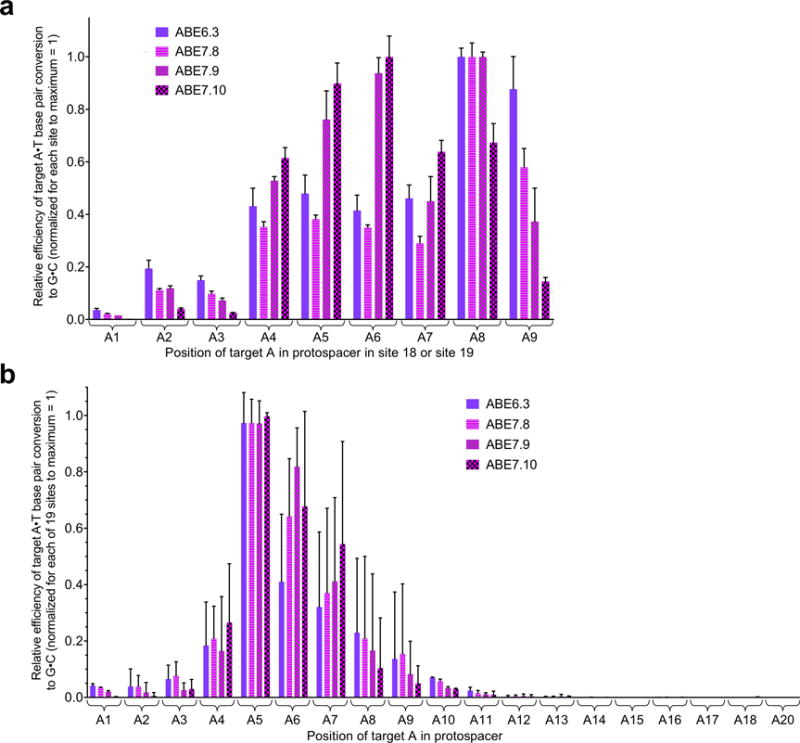
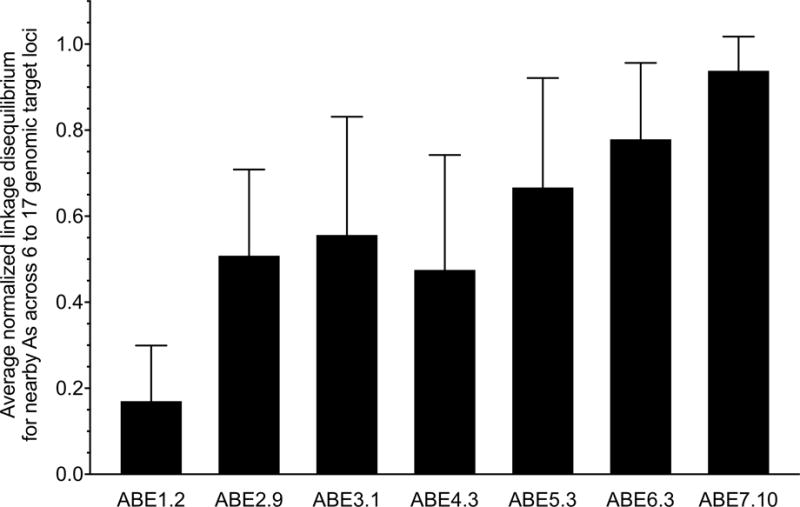
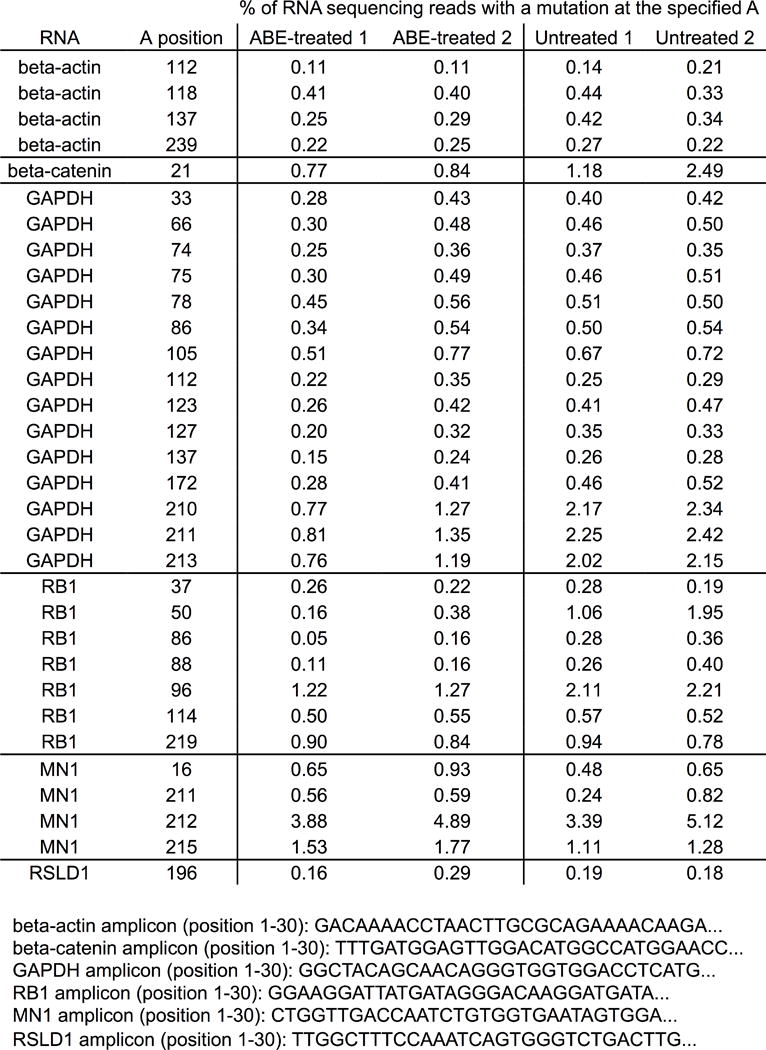
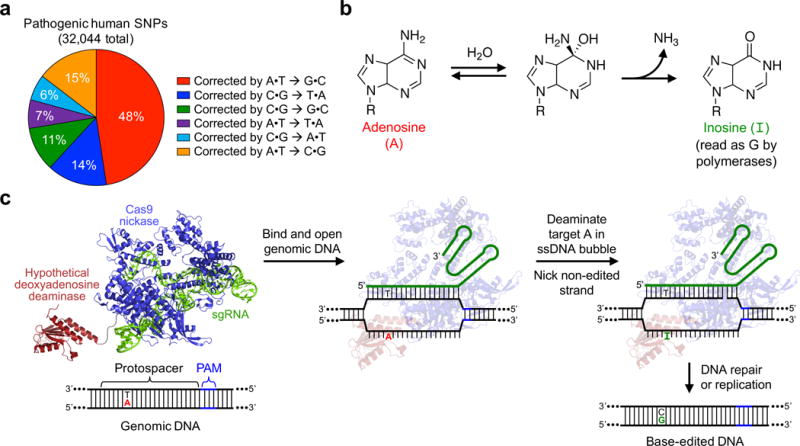
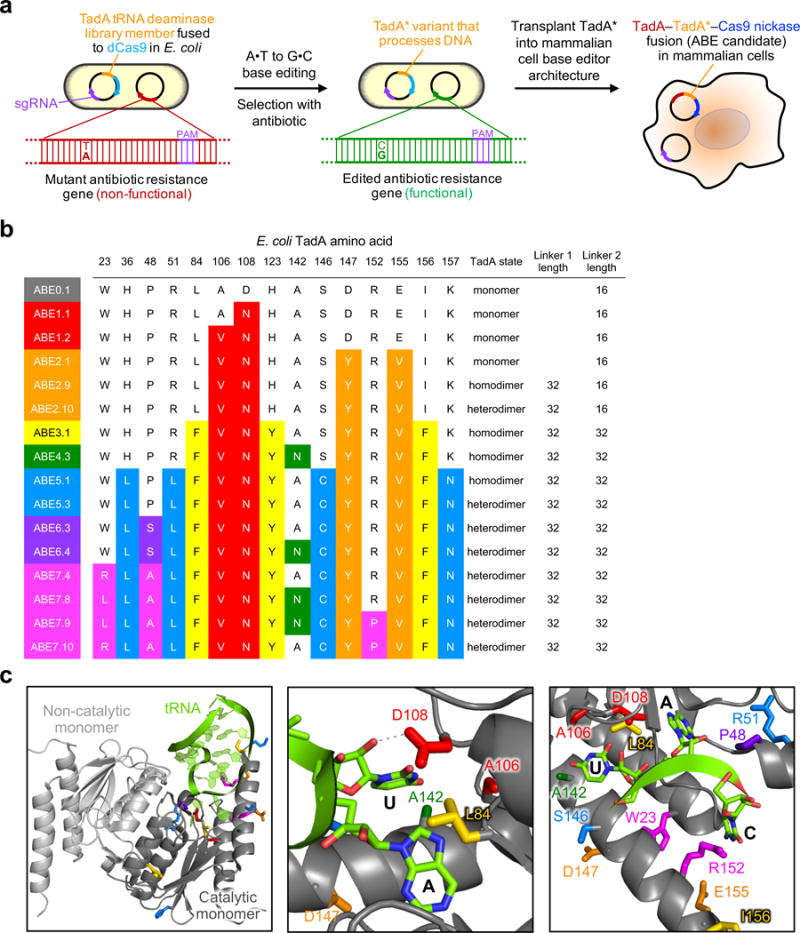
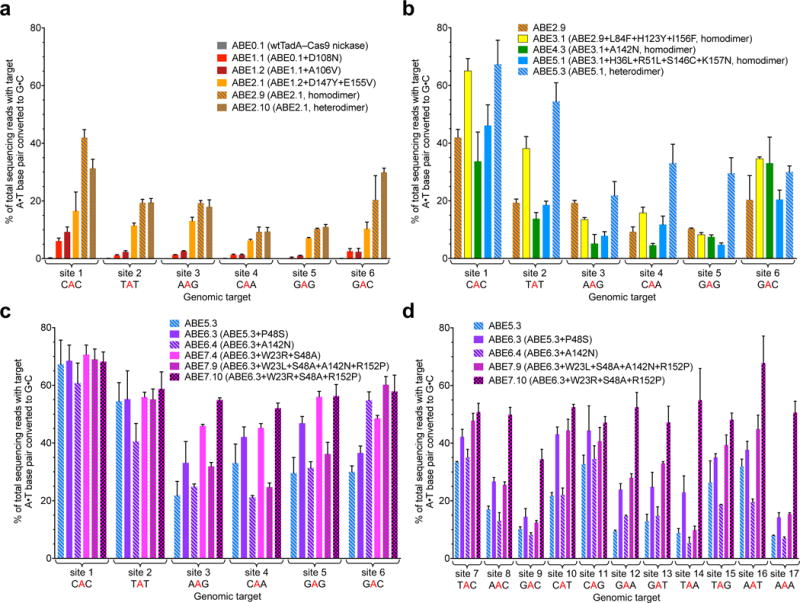
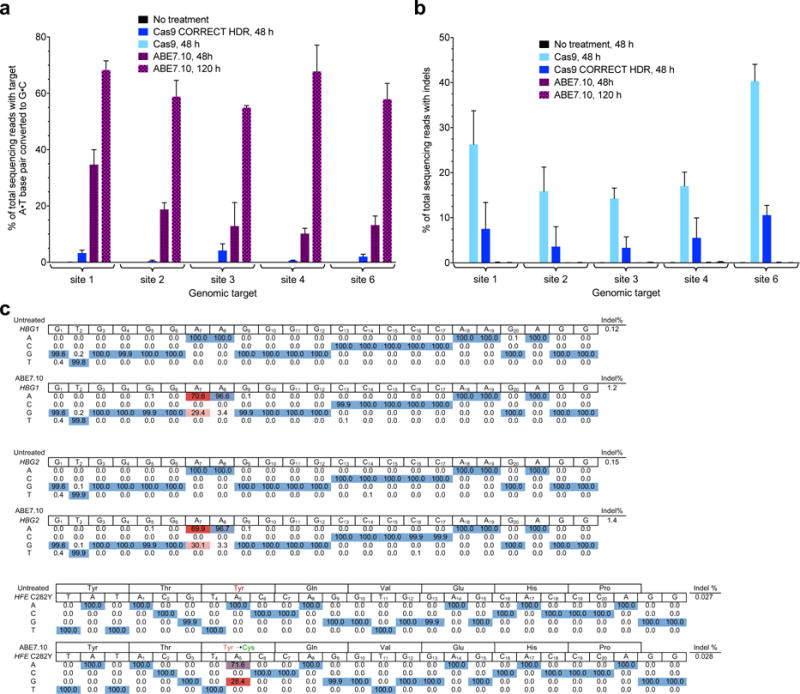
The spontaneous deamination of cytosine is a major source of transitions from C•G to T•A base pairs, which account for half of known pathogenic point mutations in humans. The ability to efficiently convert targeted A•T base pairs to G•C could therefore advance the study and treatment of genetic diseases. The deamination of adenine yields inosine, which is treated as guanine by polymerases, but no enzymes are known to deaminate adenine in DNA. Here we describe adenine base editors (ABEs) that mediate the conversion of A•T to G•C in genomic DNA. We evolved a transfer RNA adenosine deaminase to operate on DNA when fused to a catalytically impaired CRISPR-Cas9 mutant. Extensive directed evolution and protein engineering resulted in seventh-generation ABEs that convert targeted A•T base pairs efficiently to G•C (approximately 50% efficiency in human cells) with high product purity (typically at least 99.9%) and low rates of indels (typically no more than 0.1%). ABEs introduce point mutations more efficiently and cleanly, and with less off-target genome modification, than a current Cas9 nuclease-based method, and can install disease-correcting or disease-suppressing mutations in human cells. Together with previous base editors, ABEs enable the direct, programmable introduction of all four transition mutations without double-stranded DNA cleavage.
Conflict of interest statement
The authors declare competing financial interests: N.M.G., A.C.K., and D.R.L. have filed patent applications on this work. D.R.L. is a consultant and co-founder of Editas Medicine, Beam Therapeutics, and Pairwise Plants, companies that use genome editing technologies. Readers are welcome to comment on the online version of the paper.
Figures

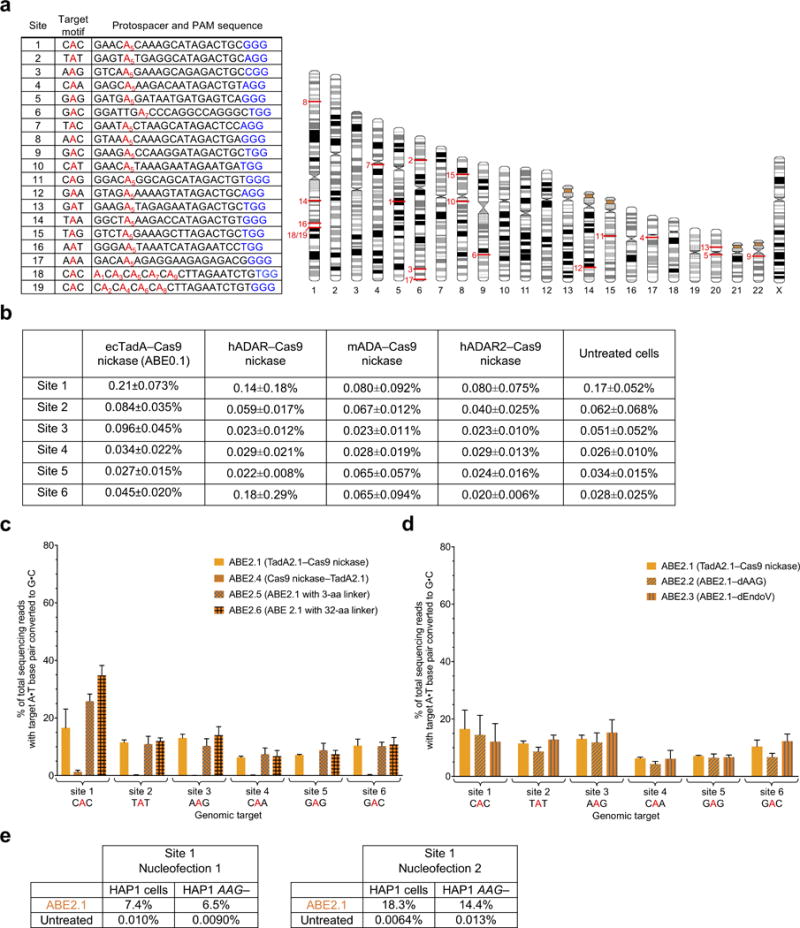
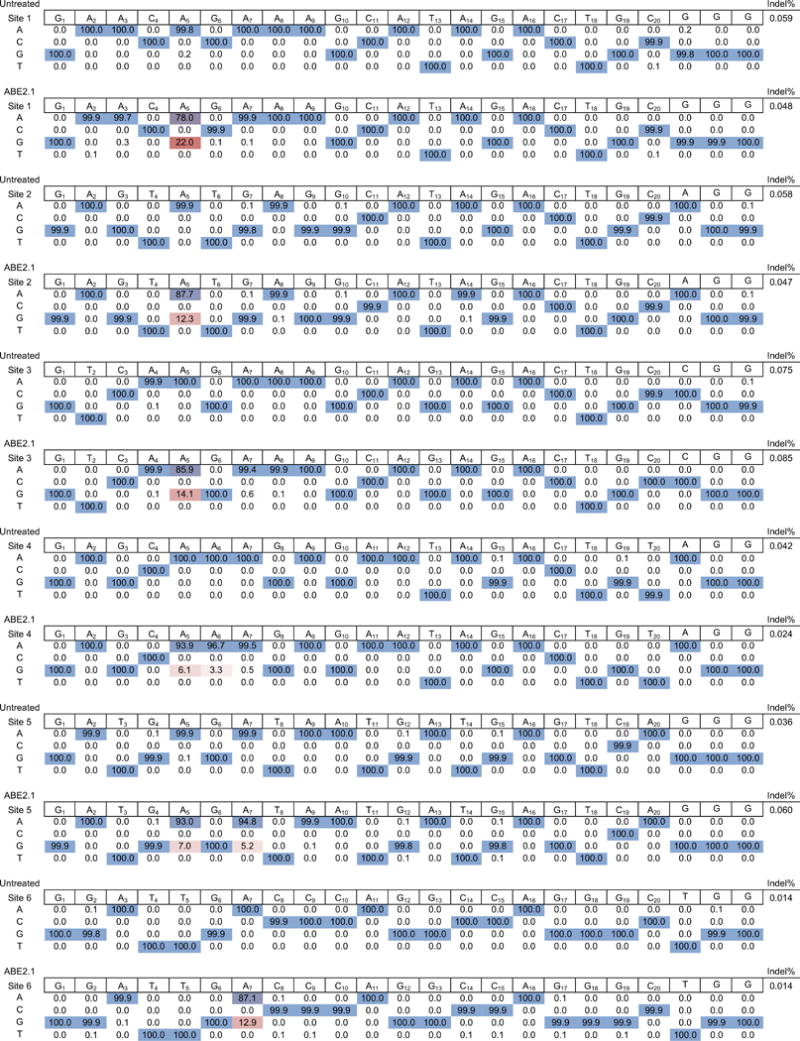
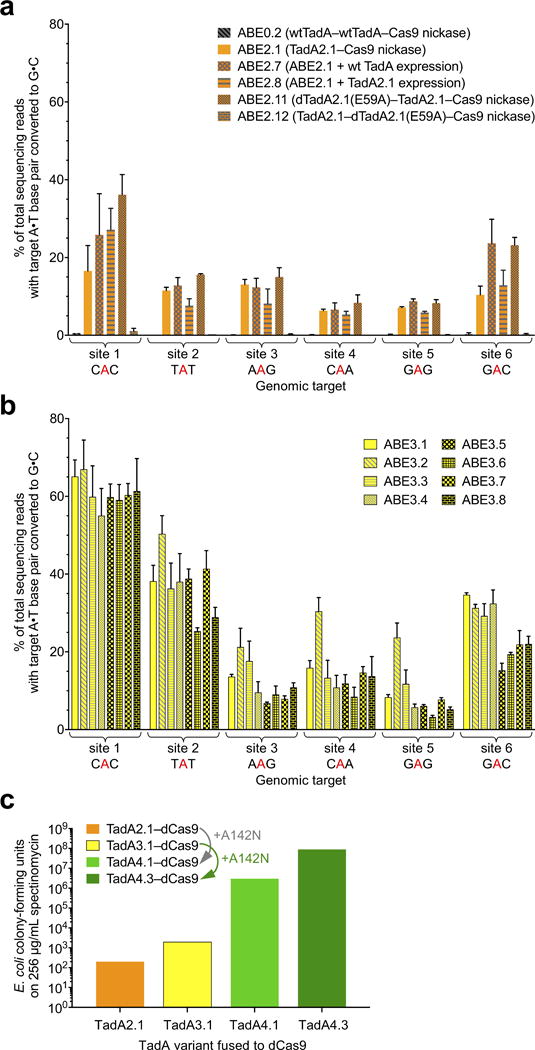
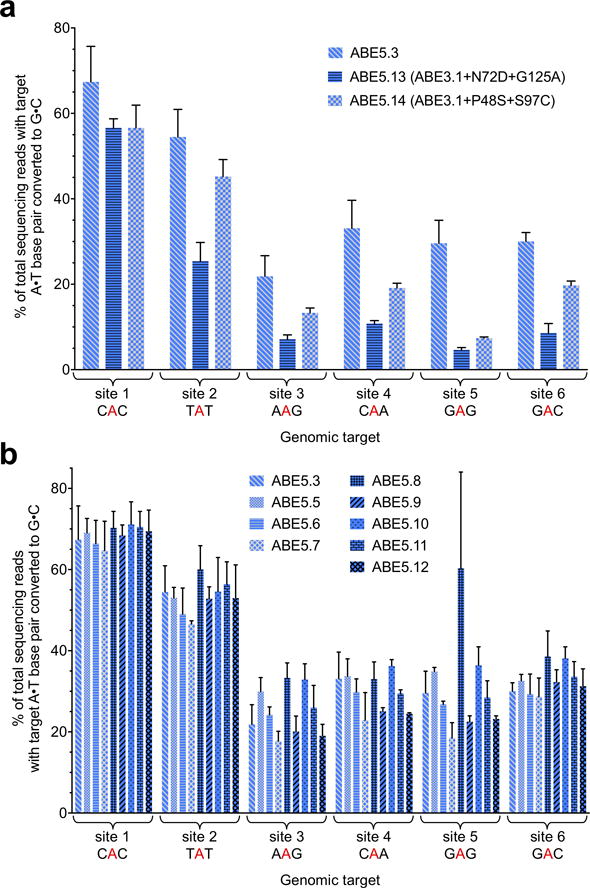


Comment in
-
CRISPR hacks enable pinpoint repairs to genome.Nature. 2017 Oct 25;550(7677):439-440. doi: 10.1038/550439a. Nature. 2017. PMID: 29072279 No abstract available.
-
Commentary: Programmable base editing of A·T to G·C in genomic DNA without DNA cleavage.Front Genet. 2018 Feb 7;9:21. doi: 10.3389/fgene.2018.00021. eCollection 2018. Front Genet. 2018. PMID: 29469899 Free PMC article. No abstract available.
References
-
- Nishida K, et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science. 2016;353 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
