

B Cells Producing Type I IFN Modulate Macrophage Polarization in Tuberculosis
- PMID: 29161093
- PMCID: PMC5855072
- DOI: 10.1164/rccm.201707-1475OC
B Cells Producing Type I IFN Modulate Macrophage Polarization in Tuberculosis
Abstract
Rationale: In addition to their well-known function as antibody-producing cells, B lymphocytes can markedly influence the course of infectious or noninfectious diseases via antibody-independent mechanisms. In tuberculosis (TB), B cells accumulate in lungs, yet their functional contribution to the host response remains poorly understood.
Objectives: To document the role of B cells in TB in an unbiased manner.
Methods: We generated the transcriptome of B cells isolated from Mycobacterium tuberculosis (Mtb)-infected mice and validated the identified key pathways using in vitro and in vivo assays. The obtained data were substantiated using B cells from pleural effusion of patients with TB.
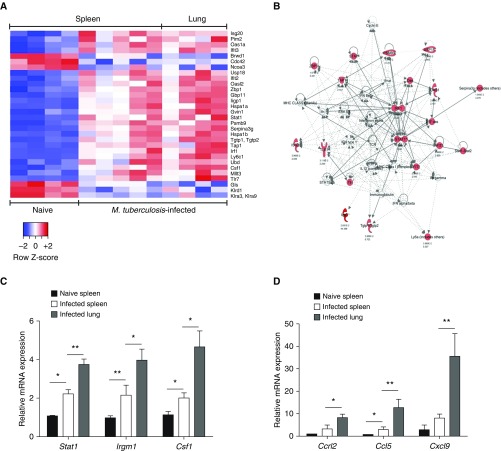
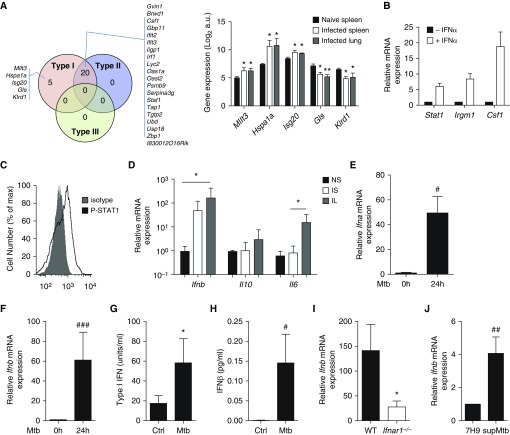
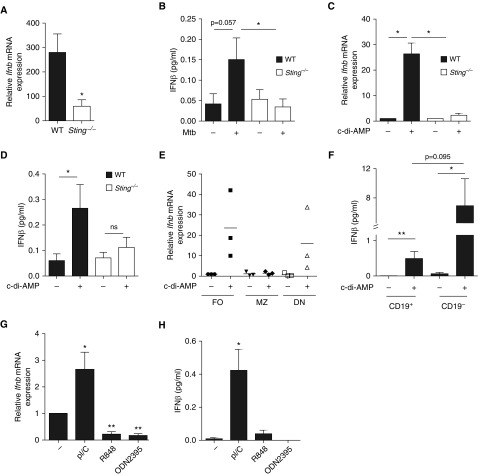
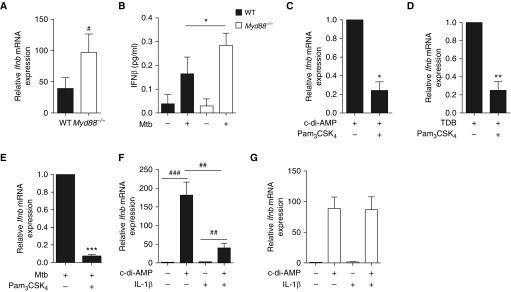
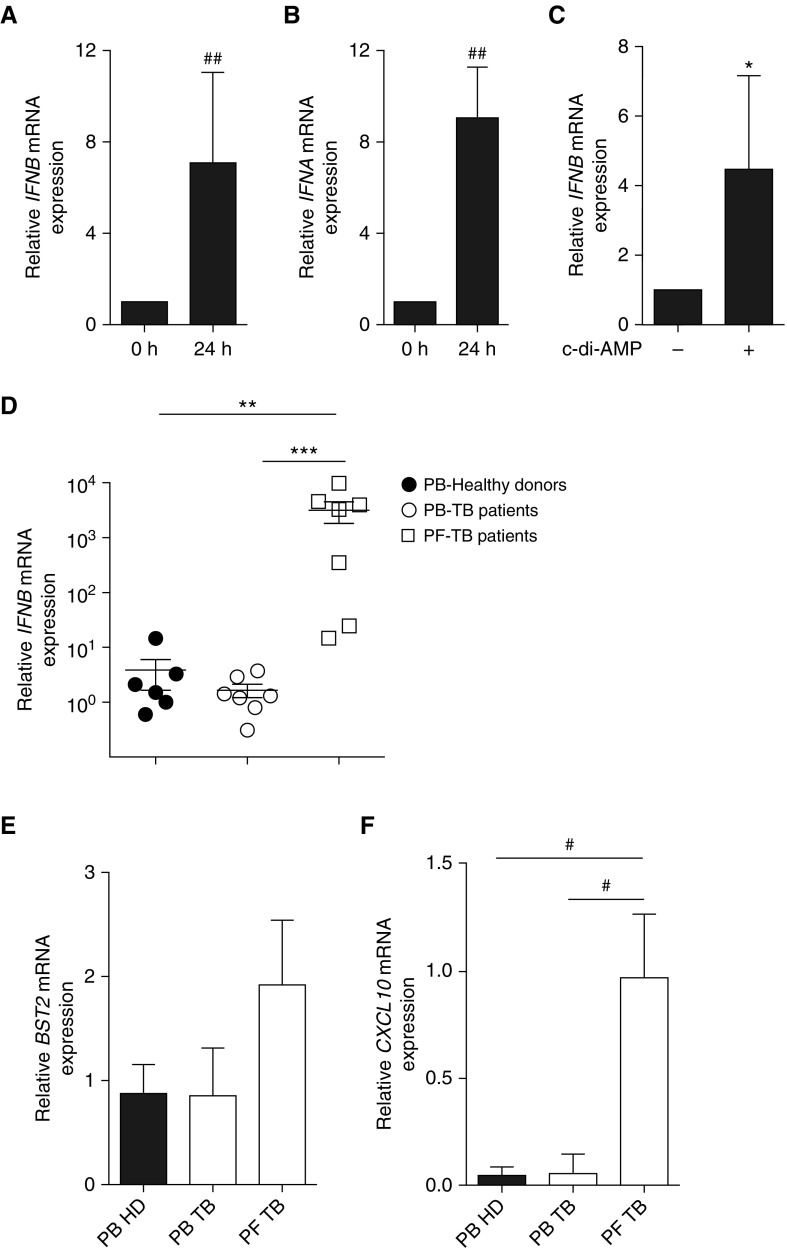
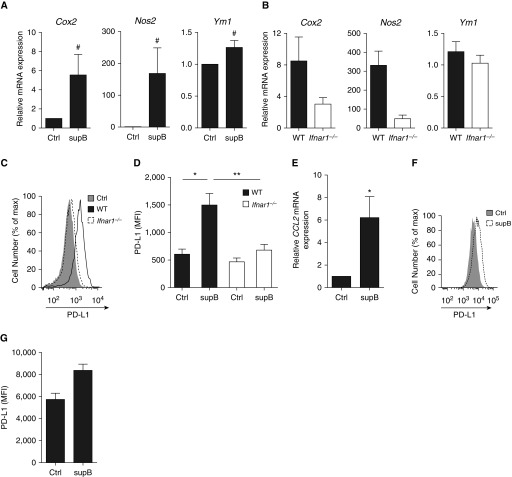
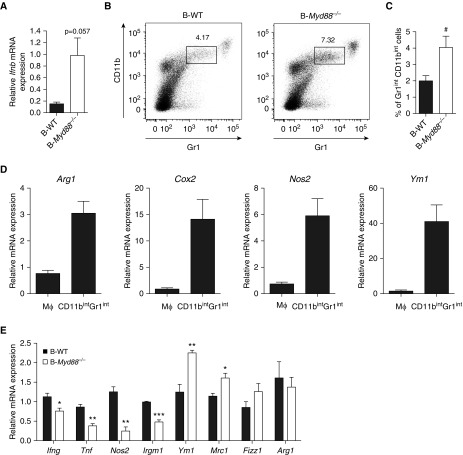
Measurements and main results: B cells isolated from Mtb-infected mice displayed a STAT1 (signal transducer and activator of transcription 1)-centered signature, suggesting a role for IFNs in B-cell response to infection. B cells stimulated in vitro with Mtb produced type I IFN, via a mechanism involving the innate sensor STING (stimulator of interferon genes), and antagonized by MyD88 (myeloid differentiation primary response 88) signaling. In vivo, B cells expressed type I IFN in the lungs of Mtb-infected mice and, of clinical relevance, in pleural fluid from patients with TB. Type I IFN expression by B cells induced an altered polarization of macrophages toward a regulatory/antiinflammatory profile in vitro. In vivo, increased provision of type I IFN by B cells in a murine model of B cell-restricted Myd88 deficiency correlated with an enhanced accumulation of regulatory/antiinflammatory macrophages in Mtb-infected lungs.
Conclusions: Type I IFN produced by Mtb-stimulated B cells favors macrophage polarization toward a regulatory/antiinflammatory phenotype during Mtb infection.
Keywords: B lymphocytes; IFN; macrophages; tuberculosis.
Figures
Comment in
-
Float Like Bacilli, STING Like a B: Type I Interferons in Tuberculosis.Am J Respir Crit Care Med. 2018 Mar 15;197(6):706-707. doi: 10.1164/rccm.201712-2368ED. Am J Respir Crit Care Med. 2018. PMID: 29262261 Free PMC article. No abstract available.
References
-
- Kahnert A, Höpken UE, Stein M, Bandermann S, Lipp M, Kaufmann SH. Mycobacterium tuberculosis triggers formation of lymphoid structure in murine lungs. J Infect Dis. 2007;195:46–54. - PubMed
-
- Maglione PJ, Xu J, Chan J. B cells moderate inflammatory progression and enhance bacterial containment upon pulmonary challenge with Mycobacterium tuberculosis. J Immunol. 2007;178:7222–7234. - PubMed
-
- Tsai MC, Chakravarty S, Zhu G, Xu J, Tanaka K, Koch C, et al. Characterization of the tuberculous granuloma in murine and human lungs: cellular composition and relative tissue oxygen tension. Cell Microbiol. 2006;8:218–232. - PubMed
-
- Ulrichs T, Kosmiadi GA, Trusov V, Jörg S, Pradl L, Titukhina M, et al. Human tuberculous granulomas induce peripheral lymphoid follicle-like structures to orchestrate local host defence in the lung. J Pathol. 2004;204:217–228. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
