

Quantitative Proteomic Profiling Reveals Novel Plasmodium falciparum Surface Antigens and Possible Vaccine Candidates
- PMID: 29162636
- PMCID: PMC5750850
- DOI: 10.1074/mcp.RA117.000076
Quantitative Proteomic Profiling Reveals Novel Plasmodium falciparum Surface Antigens and Possible Vaccine Candidates
Abstract
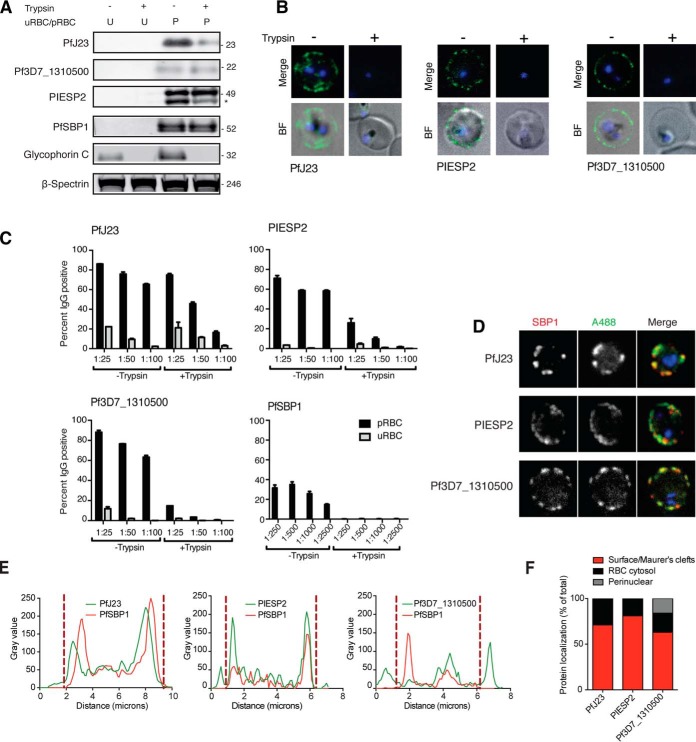
Despite recent efforts toward control and elimination, malaria remains a major public health problem worldwide. Plasmodium falciparum resistance against artemisinin, used in front line combination drugs, is on the rise, and the only approved vaccine shows limited efficacy. Combinations of novel and tailored drug and vaccine interventions are required to maintain the momentum of the current malaria elimination program. Current evidence suggests that strain-transcendent protection against malaria infection can be achieved using whole organism vaccination or with a polyvalent vaccine covering multiple antigens or epitopes. These approaches have been successfully applied to the human-infective sporozoite stage. Both systemic and tissue-specific pathology during infection with the human malaria parasite P. falciparum is caused by asexual blood stages. Tissue tropism and vascular sequestration are the result of specific binding interactions between antigens on the parasite-infected red blood cell (pRBC) surface and endothelial receptors. The major surface antigen and parasite ligand binding to endothelial receptors, PfEMP1 is encoded by about 60 variants per genome and shows high sequence diversity across strains. Apart from PfEMP1 and three additional variant surface antigen families RIFIN, STEVOR, and SURFIN, systematic analysis of the infected red blood cell surface is lacking. Here we present the most comprehensive proteomic investigation of the parasitized red blood cell surface so far. Apart from the known variant surface antigens, we identified a set of putative single copy surface antigens with low sequence diversity, several of which are validated in a series of complementary experiments. Further functional and immunological investigation is underway to test these novel P. falciparum blood stage proteins as possible vaccine candidates.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
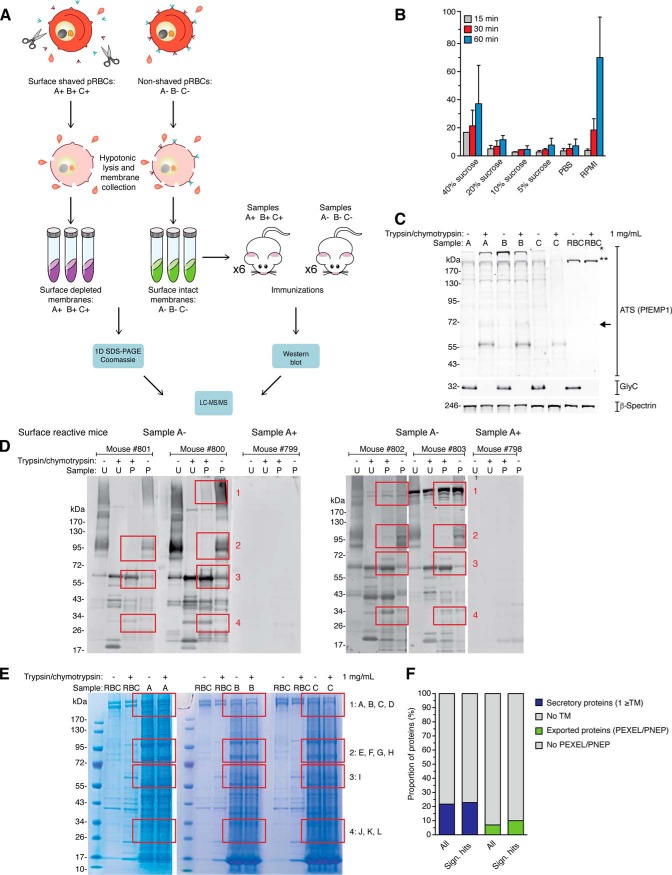
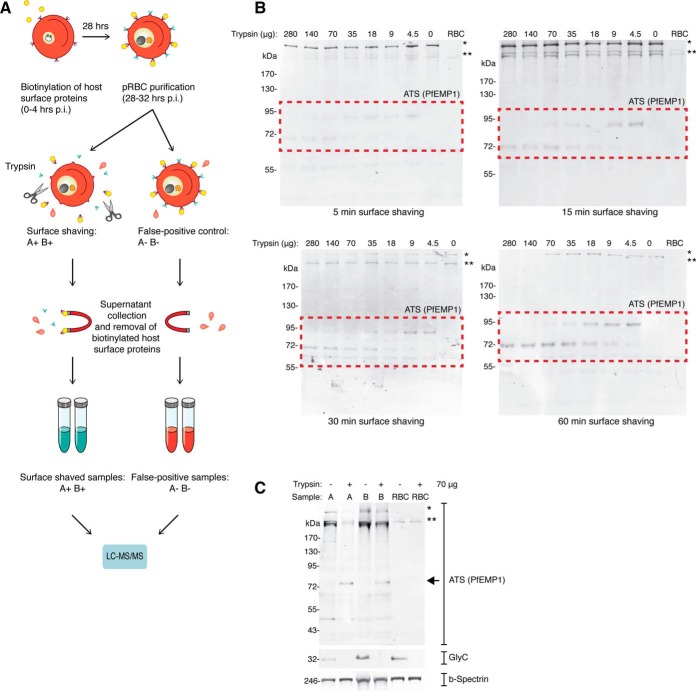
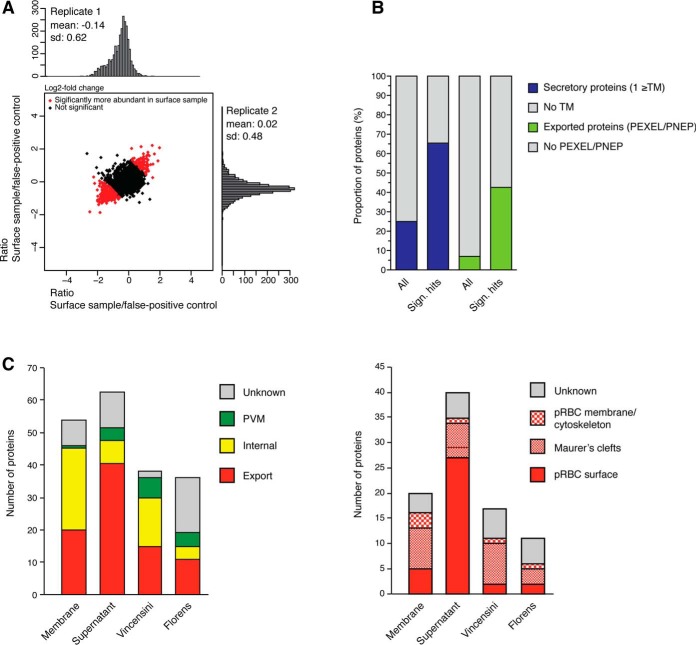
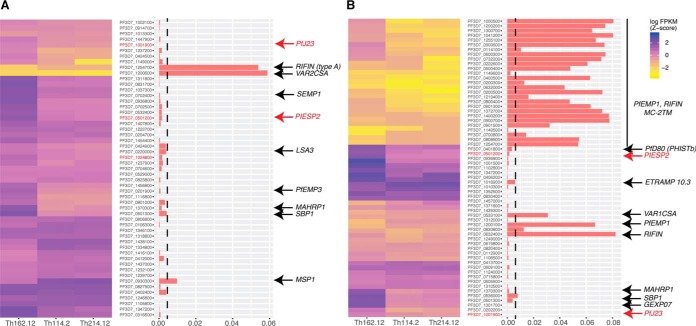
References
-
- WHO. (2016) WHO Malaria Report 2016
-
- Maier A. G., Cooke B. M., Cowman A. F., and Tilley L. (2009) Malaria parasite proteins that remodel the host erythrocyte. Nature Rev. Microbiology 7, 341–354 - PubMed
-
- Su X. Z., Heatwole V. M., Wertheimer S. P., Guinet F., Herrfeldt J. A., Peterson D. S., Ravetch J. A., and Wellems T. E. (1995) The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum-infected erythrocytes. Cell 82, 89–100 - PubMed
-
- Smith J. D., Chitnis C. E., Craig A. G., Roberts D. J., Hudson-Taylor D. E., Peterson D. S., Pinches R., Newbold C. I., and Miller L. H. (1995) Switches in expression of Plasmodium falciparum var genes correlate with changes in antigenic and cytoadherent phenotypes of infected erythrocytes. Cell 82, 101–110 - PMC - PubMed
-
- Baruch D. I., Pasloske B. L., Singh H. B., Bi X., Ma X. C., Feldman M., Taraschi T. F., and Howard R. J. (1995) Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell 82, 77–87 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
