

Hepatitis E virus: advances and challenges
- PMID: 29162935
- PMCID: PMC11329273
- DOI: 10.1038/nrgastro.2017.150
Hepatitis E virus: advances and challenges
Abstract
At least 20 million hepatitis E virus (HEV) infections occur annually, with >3 million symptomatic cases and ∼60,000 fatalities. Hepatitis E is generally self-limiting, with a case fatality rate of 0.5-3% in young adults. However, it can cause up to 30% mortality in pregnant women in the third trimester and can become chronic in immunocompromised individuals, such as those receiving organ transplants or chemotherapy and individuals with HIV infection. HEV is transmitted primarily via the faecal-oral route and was previously thought to be a public health concern only in developing countries. It is now also being frequently reported in industrialized countries, where it is transmitted zoonotically or through organ transplantation or blood transfusions. Although a vaccine for HEV has been developed, it is only licensed in China. Additionally, no effective, non-teratogenic and specific treatments against HEV infections are currently available. Although progress has been made in characterizing HEV biology, the scarcity of adequate experimental platforms has hampered further research. In this Review, we focus on providing an update on the HEV life cycle. We will further discuss existing cell culture and animal models and highlight platforms that have proven to be useful and/or are emerging for studying other hepatotropic (viral) pathogens.
Conflict of interest statement
Competing interests statement
A.P. and Q.D. are inventors on a patent application defining an hepatitis E virus transcomplementation system for antiviral drug screening and ORF3’s viroporin function as a antiviral drug target.
Figures
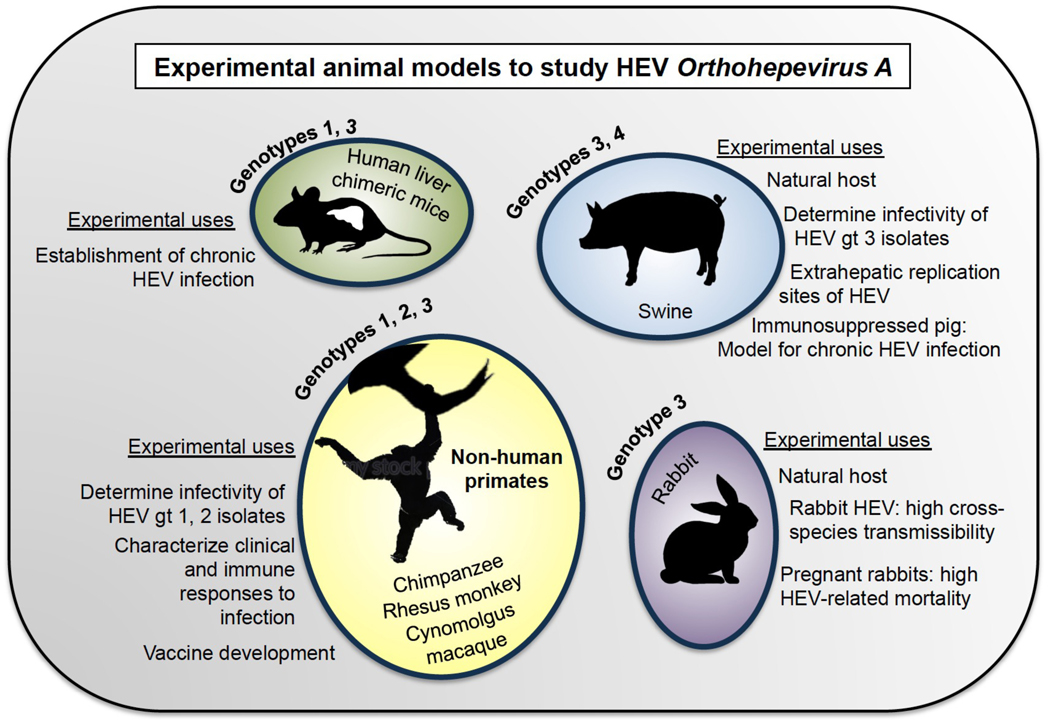
References
-
- Khuroo MS Study of an epidemic of non-A, non-B hepatitis. Possibility of another human hepatitis virus distinct from post-transfusion non-A, non-B type. Am J Med 68, 818–824 (1980). - PubMed
-
- Wong DC, Purcell RH, Sreenivasan MA, Prasad SR & Pavri KM Epidemic and endemic hepatitis in India: evidence for a non-A, non-B hepatitis virus aetiology. Lancet 2, 876–879 (1980). - PubMed
-
-
Balayan MS et al. Evidence for a virus in non-A, non-B hepatitis transmitted via the fecal-oral route. Intervirology 20, 23–31 (1983).
A volunteer ingested pooled stool extracts from patients infected with HEV to show for the first time that the virus is fecal-orally transmitted.
-
-
-
Reyes GR et al. Isolation of a cDNA from the virus responsible for enterically transmitted nonA, non-B hepatitis. Science 247, 1335–1339 (1990).
The first partial cDNA of ET-NANBH was cloned, and the virus renamed ‘hepatitis E virus’.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
