

Aerobic performance in tinamous is limited by their small heart. A novel hypothesis in the evolution of avian flight
- PMID: 29162941
- PMCID: PMC5698454
- DOI: 10.1038/s41598-017-16297-2
Aerobic performance in tinamous is limited by their small heart. A novel hypothesis in the evolution of avian flight
Abstract
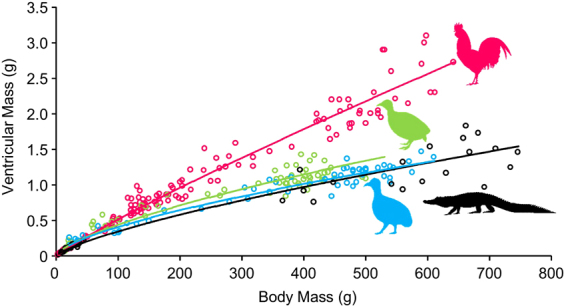
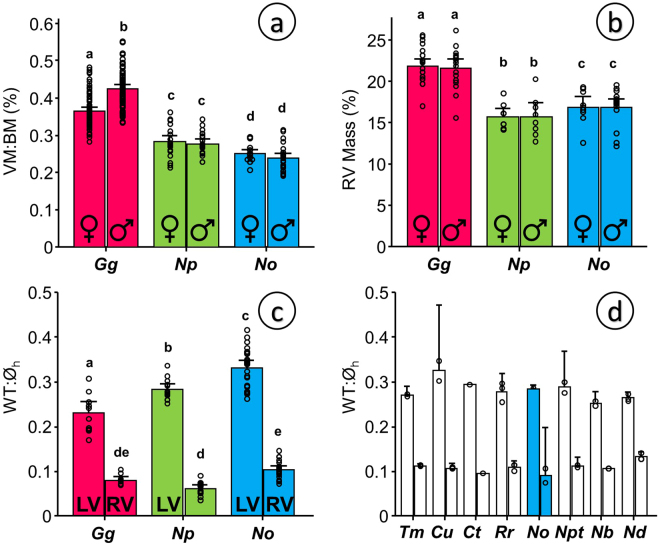
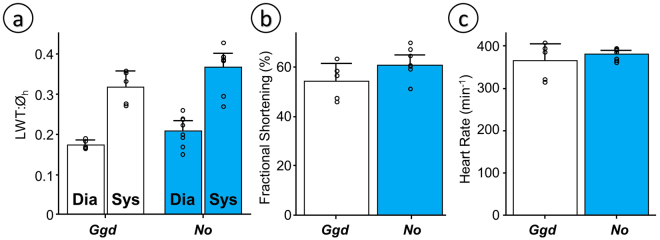
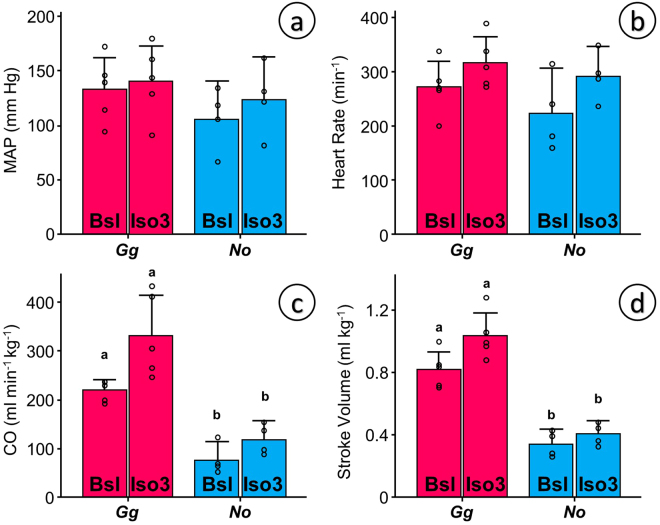
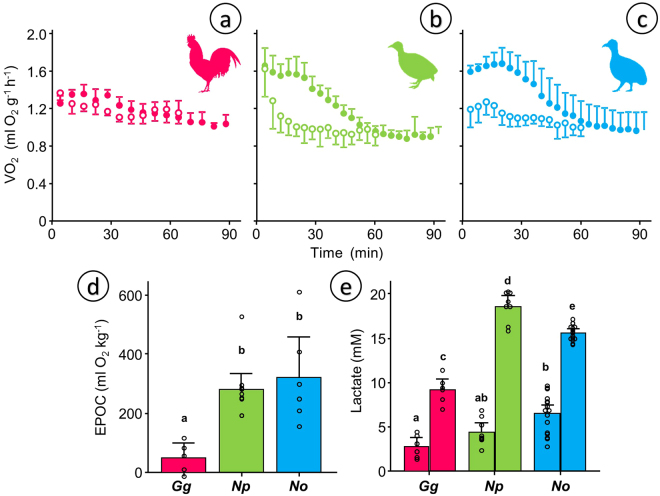
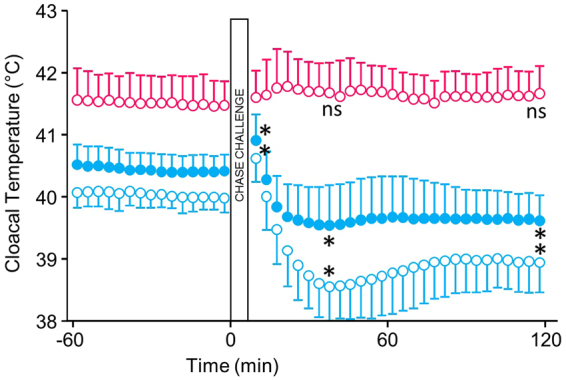
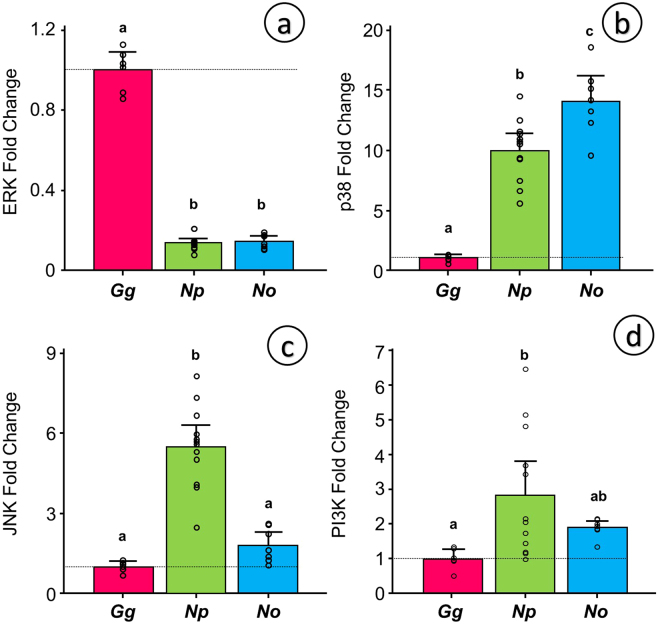
Some biomechanical studies from fossil specimens suggest that sustained flapping flight of birds could have appeared in their Mesozoic ancestors. We challenge this idea because a suitable musculoskeletal anatomy is not the only requirement for sustained flapping flight. We propose the "heart to fly" hypothesis that states that sustained flapping flight in modern birds required an enlargement of the heart for the aerobic performance of the flight muscles and test it experimentally by studying tinamous, the living birds with the smallest hearts. The small ventricular size of tinamous reduces cardiac output without limiting perfusion pressures, but when challenged to fly, the heart is unable to support aerobic metabolism (quick exhaustion, larger lactates and post-exercise oxygen consumption and compromised thermoregulation). At the same time, cardiac growth shows a crocodilian-like pattern and is correlated with differential gene expression in MAPK kinases. We integrate this physiological evidence in a new evolutionary scenario in which the ground-up, short and not sustained flapping flight displayed by tinamous represents an intermediate step in the evolution of the aerobic sustained flapping flight of modern birds.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Feduccia A. Bird origins anew. The Auk. 2013;130:1–12. doi: 10.1525/auk.2013.130.1.1. - DOI
-
- Smith NA, et al. Rhetoric vs. reality: a commentary on “Bird Origins Anew” by A.Feduccia. The Auk. 2015;132:467–480. doi: 10.1642/AUK-14-203.1. - DOI
-
- Padian K, Chiappe LM. The origin and early evolution of birds. Biol.Rev. 1998;73:1–42. doi: 10.1017/S0006323197005100. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
