

Functional 5' UTR mRNA structures in eukaryotic translation regulation and how to find them
- PMID: 29165424
- PMCID: PMC5820134
- DOI: 10.1038/nrm.2017.103
Functional 5' UTR mRNA structures in eukaryotic translation regulation and how to find them
Erratum in
-
Author Correction: Functional 5' UTR mRNA structures in eukaryotic translation regulation and how to find them.Nat Rev Mol Cell Biol. 2018 Oct;19(10):673. doi: 10.1038/s41580-018-0055-5. Nat Rev Mol Cell Biol. 2018. PMID: 30111874
Abstract
RNA molecules can fold into intricate shapes that can provide an additional layer of control of gene expression beyond that of their sequence. In this Review, we discuss the current mechanistic understanding of structures in 5' untranslated regions (UTRs) of eukaryotic mRNAs and the emerging methodologies used to explore them. These structures may regulate cap-dependent translation initiation through helicase-mediated remodelling of RNA structures and higher-order RNA interactions, as well as cap-independent translation initiation through internal ribosome entry sites (IRESs), mRNA modifications and other specialized translation pathways. We discuss known 5' UTR RNA structures and how new structure probing technologies coupled with prospective validation, particularly compensatory mutagenesis, are likely to identify classes of structured RNA elements that shape post-transcriptional control of gene expression and the development of multicellular organisms.
Conflict of interest statement
The authors declare no competing interests.
Figures
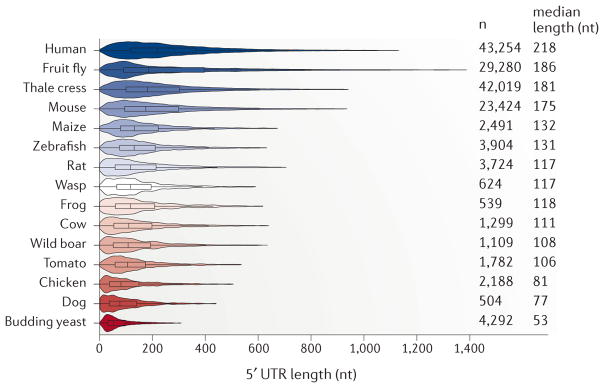
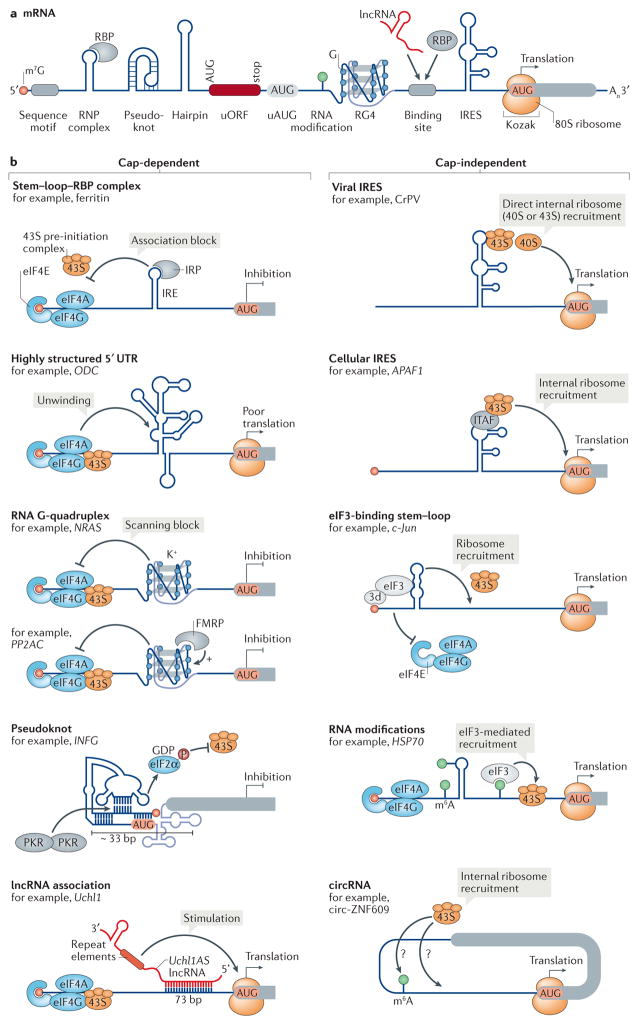
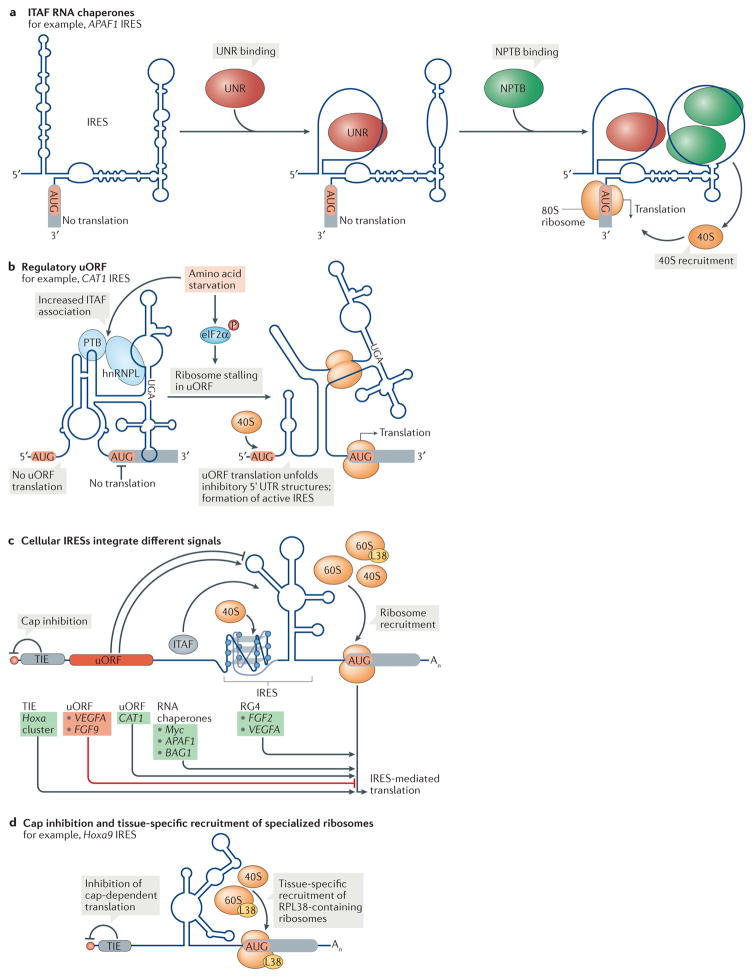
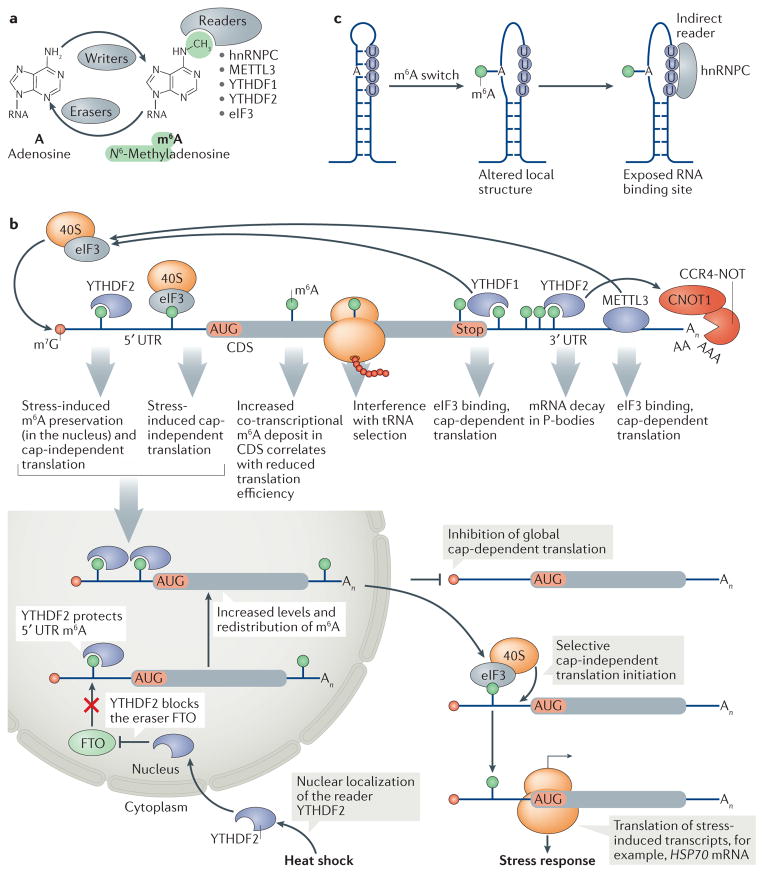
References
-
- Bowman JC, Hud NV, Williams LD. The ribosome challenge to the RNA world. J Mol Evol. 2015;80:143–161. - PubMed
-
- Nahvi A, et al. Genetic control by a metabolite binding mRNA. Chem Biol. 2002;9:1043–1049. - PubMed
-
- Wachter A. Gene regulation by structured mRNA elements. Trends Genet. 2014;30:172–181. - PubMed
-
- Martin W, Koonin EV. Introns and the origin of nucleus-cytosol compartmentalization. Nature. 2006;440:41–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
