

Granulocyte-Monocyte Progenitors and Monocyte-Dendritic Cell Progenitors Independently Produce Functionally Distinct Monocytes
- PMID: 29166589
- PMCID: PMC5726802
- DOI: 10.1016/j.immuni.2017.10.021
Granulocyte-Monocyte Progenitors and Monocyte-Dendritic Cell Progenitors Independently Produce Functionally Distinct Monocytes
Abstract
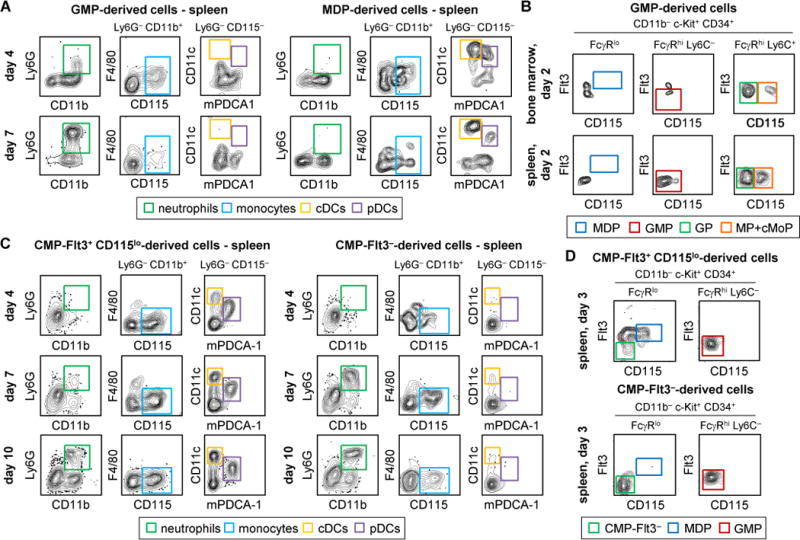
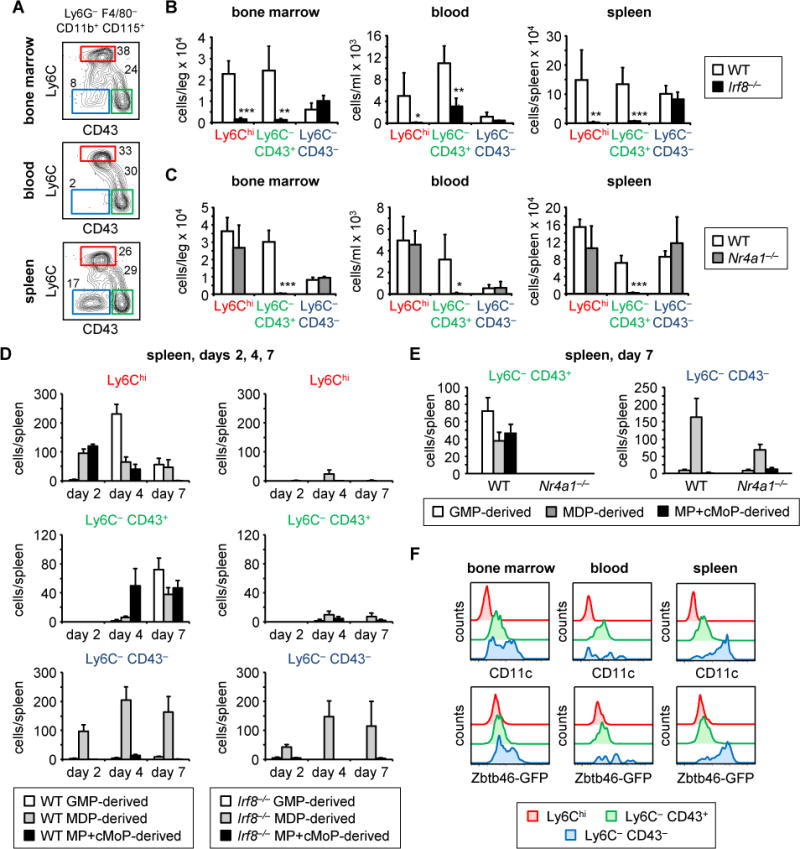
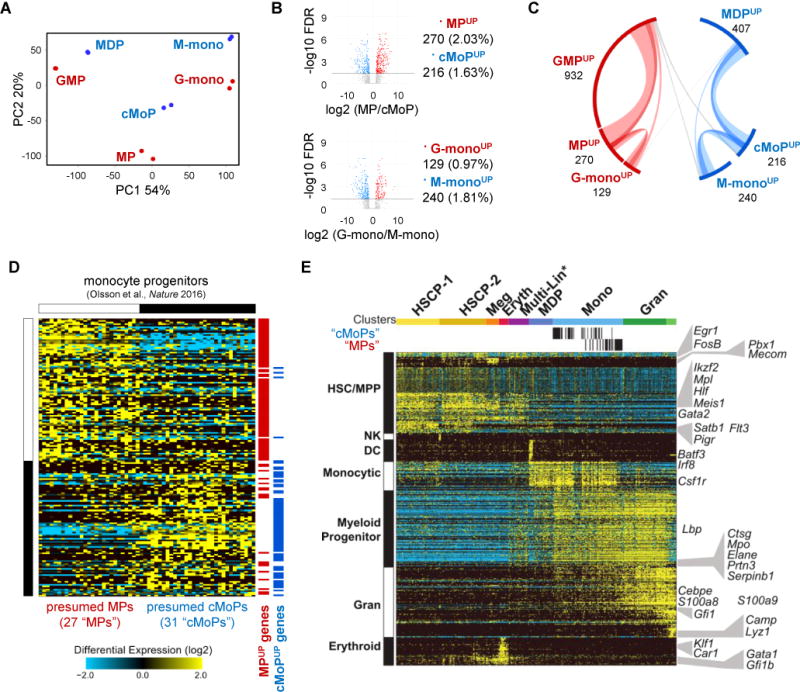
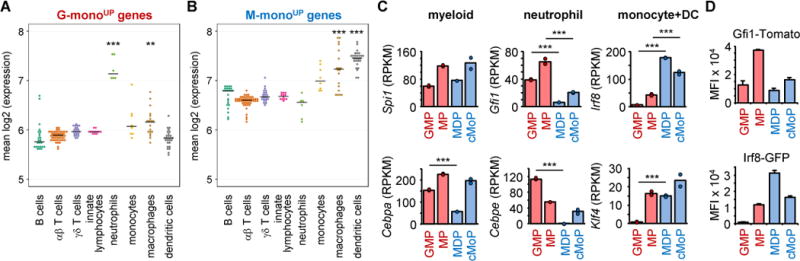
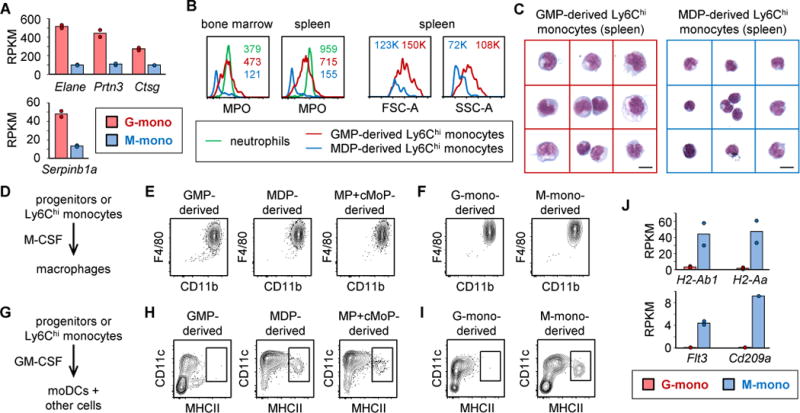
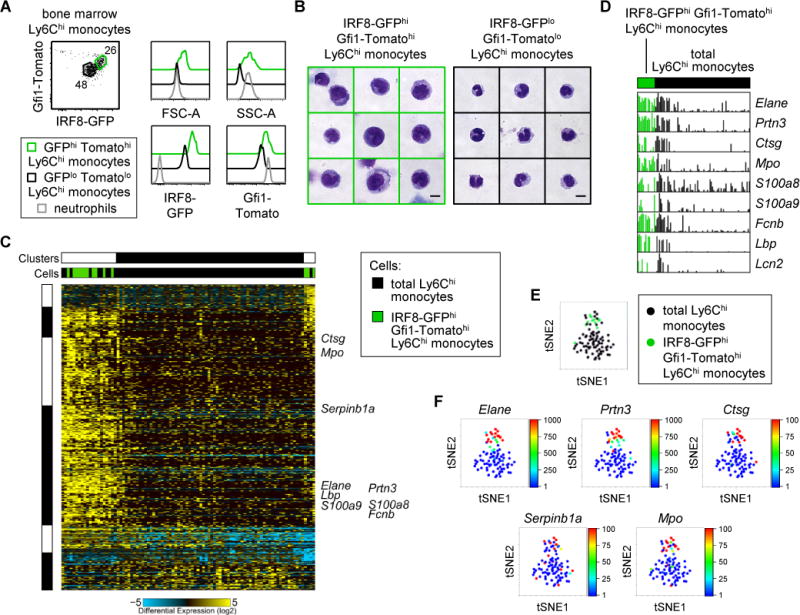
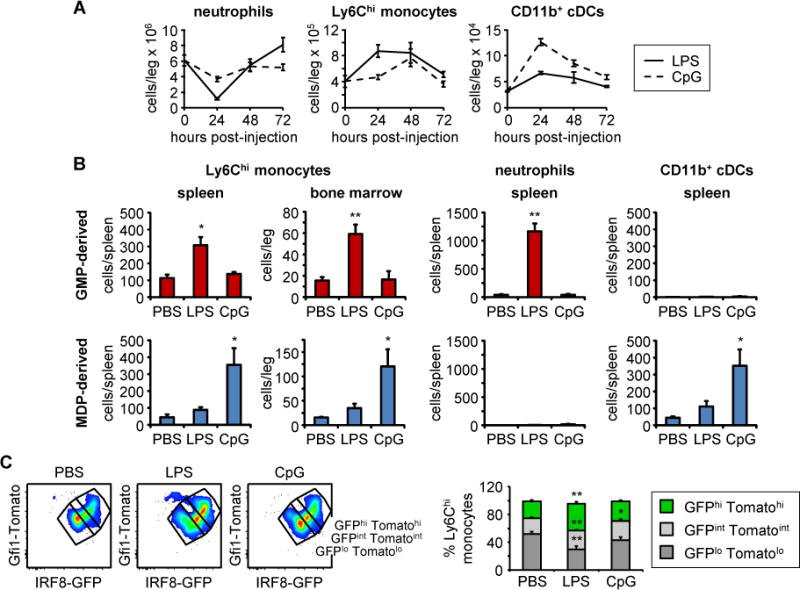
Granulocyte-monocyte progenitors (GMPs) and monocyte-dendritic cell progenitors (MDPs) produce monocytes during homeostasis and in response to increased demand during infection. Both progenitor populations are thought to derive from common myeloid progenitors (CMPs), and a hierarchical relationship (CMP-GMP-MDP-monocyte) is presumed to underlie monocyte differentiation. Here, however, we demonstrate that mouse MDPs arose from CMPs independently of GMPs, and that GMPs and MDPs produced monocytes via similar but distinct monocyte-committed progenitors. GMPs and MDPs yielded classical (Ly6Chi) monocytes with gene expression signatures that were defined by their origins and impacted their function. GMPs produced a subset of "neutrophil-like" monocytes, whereas MDPs gave rise to a subset of monocytes that yielded monocyte-derived dendritic cells. GMPs and MDPs were also independently mobilized to produce specific combinations of myeloid cell types following the injection of microbial components. Thus, the balance of GMP and MDP differentiation shapes the myeloid cell repertoire during homeostasis and following infection.
Keywords: GMP; MDP; MP; cMoP; dendritic cell; granulocyte-monocyte progenitor; monocyte; monocyte progenitor; monocyte-dendritic cell progenitor; myeloid cell; myeloid progenitor; myelopoiesis; neutrophil.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Akashi K, Traver D, Miyamoto T, Weissman IL. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature. 2000;404:193–197. - PubMed
-
- Basu S, Hodgson G, Zhang HH, Katz M, Quilici C, Dunn AR. “Emergency” granulopoiesis in G-CSF-deficient mice in response to Candida albicans infection. Blood. 2000;95:3725–3733. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
