

Fragmentation modes and the evolution of life cycles
- PMID: 29166656
- PMCID: PMC5718564
- DOI: 10.1371/journal.pcbi.1005860
Fragmentation modes and the evolution of life cycles
Abstract
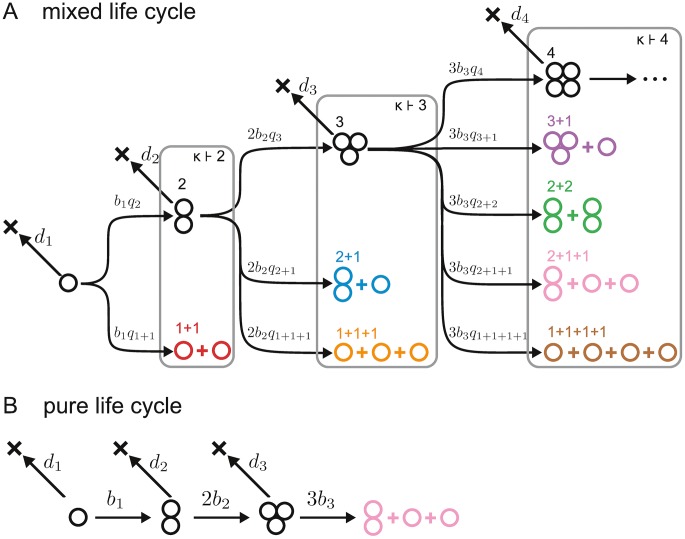
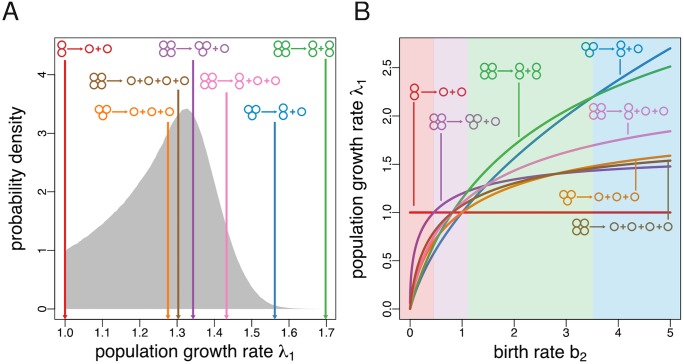
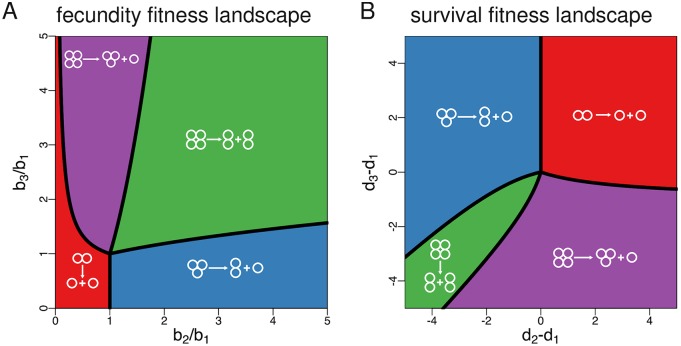
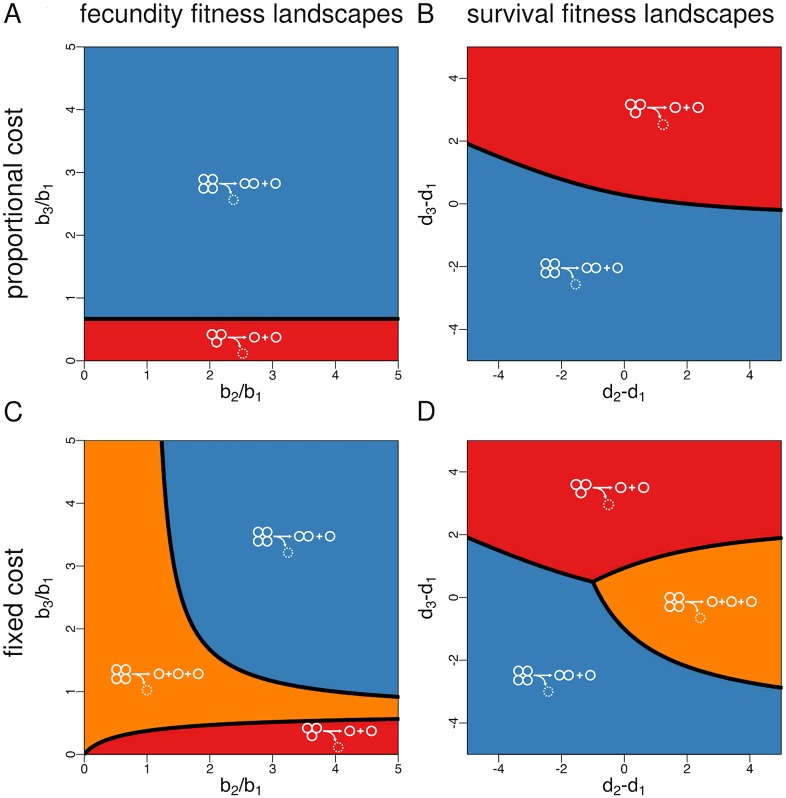
Reproduction is a defining feature of living systems. To reproduce, aggregates of biological units (e.g., multicellular organisms or colonial bacteria) must fragment into smaller parts. Fragmentation modes in nature range from binary fission in bacteria to collective-level fragmentation and the production of unicellular propagules in multicellular organisms. Despite this apparent ubiquity, the adaptive significance of fragmentation modes has received little attention. Here, we develop a model in which groups arise from the division of single cells that do not separate but stay together until the moment of group fragmentation. We allow for all possible fragmentation patterns and calculate the population growth rate of each associated life cycle. Fragmentation modes that maximise growth rate comprise a restrictive set of patterns that include production of unicellular propagules and division into two similar size groups. Life cycles marked by single-cell bottlenecks maximise population growth rate under a wide range of conditions. This surprising result offers a new evolutionary explanation for the widespread occurrence of this mode of reproduction. All in all, our model provides a framework for exploring the adaptive significance of fragmentation modes and their associated life cycles.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Interacting cells driving the evolution of multicellular life cycles.PLoS Comput Biol. 2019 May 14;15(5):e1006987. doi: 10.1371/journal.pcbi.1006987. eCollection 2019 May. PLoS Comput Biol. 2019. PMID: 31086369 Free PMC article.
-
Evolution of multicellular life cycles under costly fragmentation.PLoS Comput Biol. 2020 Nov 19;16(11):e1008406. doi: 10.1371/journal.pcbi.1008406. eCollection 2020 Nov. PLoS Comput Biol. 2020. PMID: 33211685 Free PMC article.
-
A Darwinian approach to the origin of life cycles with group properties.Theor Popul Biol. 2015 Jun;102:76-84. doi: 10.1016/j.tpb.2015.03.003. Epub 2015 Mar 23. Theor Popul Biol. 2015. PMID: 25814207
-
Evolution of complex life cycles in trophically transmitted helminths. I. Host incorporation and trophic ascent.J Evol Biol. 2015 Feb;28(2):267-91. doi: 10.1111/jeb.12575. Epub 2015 Jan 27. J Evol Biol. 2015. PMID: 25625702 Review.
-
Evolution of complex life cycles in trophically transmitted helminths. II. How do life-history stages adapt to their hosts?J Evol Biol. 2015 Feb;28(2):292-304. doi: 10.1111/jeb.12576. Epub 2015 Jan 27. J Evol Biol. 2015. PMID: 25645609 Review.
Cited by
-
Stability of ecologically scaffolded traits during evolutionary transitions in individuality.Nat Commun. 2024 Aug 3;15(1):6566. doi: 10.1038/s41467-024-50625-1. Nat Commun. 2024. PMID: 39095362 Free PMC article.
-
Evolution of reproductive strategies in incipient multicellularity.J R Soc Interface. 2022 Mar;19(188):20210716. doi: 10.1098/rsif.2021.0716. Epub 2022 Mar 2. J R Soc Interface. 2022. PMID: 35232276 Free PMC article.
-
Alternating selection for dispersal and multicellularity favors regulated life cycles.Curr Biol. 2023 May 8;33(9):1809-1817.e3. doi: 10.1016/j.cub.2023.03.031. Epub 2023 Apr 4. Curr Biol. 2023. PMID: 37019107 Free PMC article.
-
Evolution of irreversible somatic differentiation.Elife. 2021 Oct 13;10:e66711. doi: 10.7554/eLife.66711. Elife. 2021. PMID: 34643506 Free PMC article.
-
Interacting cells driving the evolution of multicellular life cycles.PLoS Comput Biol. 2019 May 14;15(5):e1006987. doi: 10.1371/journal.pcbi.1006987. eCollection 2019 May. PLoS Comput Biol. 2019. PMID: 31086369 Free PMC article.
References
-
- Godfrey-Smith P. Darwinian populations and natural selection. Oxford University Press; 2009.
-
- Libby E, Rainey PB. A conceptual framework for the evolutionary origins of multicellularity. Physical Biology. 2013;10(3):035001 doi: 10.1088/1478-3975/10/3/035001 - DOI - PubMed
-
- Hammerschmidt K, Rose CJ, Kerr B, Rainey PB. Life cycles, fitness decoupling and the evolution of multicellularity. Nature. 2014;515(7525):75–79. doi: 10.1038/nature13884 - DOI - PubMed
-
- van Gestel J, Tarnita CE. On the origin of biological construction, with a focus on multicellularity. Proceedings of the National Academy of Sciences. 2017;114(42):11018–11026. doi: 10.1073/pnas.1704631114 - DOI - PMC - PubMed
-
- Angert ER. Alternatives to binary fission in bacteria. Nature Reviews Microbiology. 2005;3(3):214–224. doi: 10.1038/nrmicro1096 - DOI - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
