

DRC2/CCDC65 is a central hub for assembly of the nexin-dynein regulatory complex and other regulators of ciliary and flagellar motility
- PMID: 29167384
- PMCID: PMC5909927
- DOI: 10.1091/mbc.E17-08-0510
DRC2/CCDC65 is a central hub for assembly of the nexin-dynein regulatory complex and other regulators of ciliary and flagellar motility
Abstract
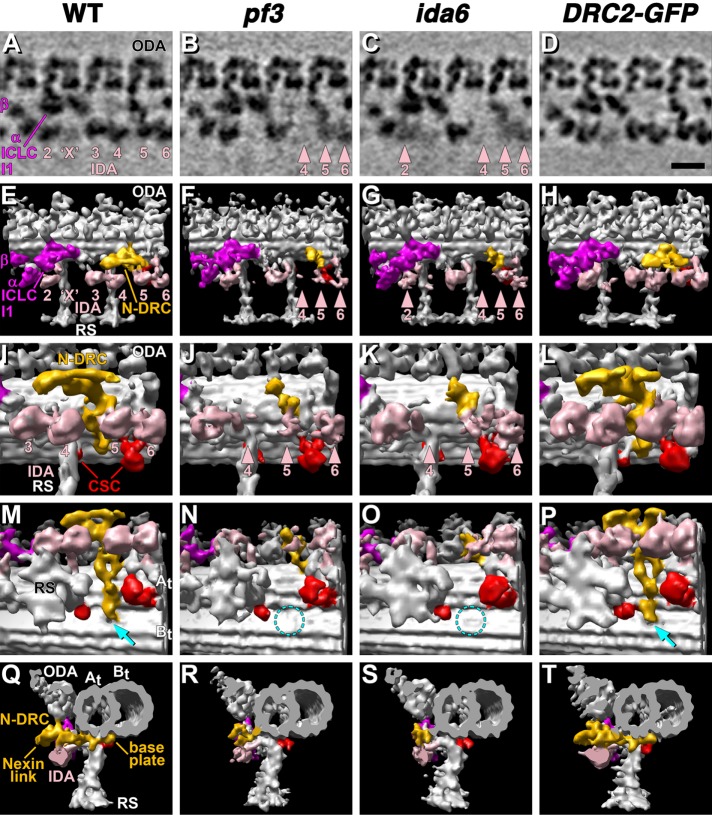
The nexin-dynein regulatory complex (N-DRC) plays a central role in the regulation of ciliary and flagellar motility. In most species, the N-DRC contains at least 11 subunits, but the specific function of each subunit is unknown. Mutations in three subunits (DRC1, DRC2/CCDC65, DRC4/GAS8) have been linked to defects in ciliary motility in humans and lead to a ciliopathy known as primary ciliary dyskinesia (PCD). Here we characterize the biochemical, structural, and motility phenotypes of two mutations in the DRC2 gene of Chlamydomonas Using high-resolution proteomic and structural approaches, we find that the C-terminal region of DRC2 is critical for the coassembly of DRC2 and DRC1 to form the base plate of N-DRC and its attachment to the outer doublet microtubule. Loss of DRC2 in drc2 mutants disrupts the assembly of several other N-DRC subunits and also destabilizes the assembly of several closely associated structures such as the inner dynein arms, the radial spokes, and the calmodulin- and spoke-associated complex. Our study provides new insights into the range of ciliary defects that can lead to PCD.
© 2018 Bower et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures






Similar articles
-
CCDC65 mutation causes primary ciliary dyskinesia with normal ultrastructure and hyperkinetic cilia.PLoS One. 2013 Aug 26;8(8):e72299. doi: 10.1371/journal.pone.0072299. eCollection 2013. PLoS One. 2013. PMID: 23991085 Free PMC article.
-
Detailed structural and biochemical characterization of the nexin-dynein regulatory complex.Mol Biol Cell. 2015 Jan 15;26(2):294-304. doi: 10.1091/mbc.E14-09-1367. Epub 2014 Nov 19. Mol Biol Cell. 2015. PMID: 25411337 Free PMC article.
-
Scaffold subunits support associated subunit assembly in the Chlamydomonas ciliary nexin-dynein regulatory complex.Proc Natl Acad Sci U S A. 2019 Nov 12;116(46):23152-23162. doi: 10.1073/pnas.1910960116. Epub 2019 Oct 28. Proc Natl Acad Sci U S A. 2019. PMID: 31659045 Free PMC article.
-
Primary Ciliary Dyskinesia.2007 Jan 24 [updated 2025 May 22]. In: Adam MP, Feldman J, Mirzaa GM, Pagon RA, Wallace SE, Amemiya A, editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2025. 2007 Jan 24 [updated 2025 May 22]. In: Adam MP, Feldman J, Mirzaa GM, Pagon RA, Wallace SE, Amemiya A, editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2025. PMID: 20301301 Free Books & Documents. Review.
-
Ciliary Motility: Regulation of Axonemal Dynein Motors.Cold Spring Harb Perspect Biol. 2017 Aug 1;9(8):a018325. doi: 10.1101/cshperspect.a018325. Cold Spring Harb Perspect Biol. 2017. PMID: 28765157 Free PMC article. Review.
Cited by
-
A Nonsense Variant in CCDC65 Gene Causes Respiratory Failure Associated with Increased Lamb Mortality in French Lacaune Dairy Sheep.Genes (Basel). 2021 Dec 24;13(1):45. doi: 10.3390/genes13010045. Genes (Basel). 2021. PMID: 35052387 Free PMC article.
-
CCDC65 as a new potential tumor suppressor induced by metformin inhibits activation of AKT1 via ubiquitination of ENO1 in gastric cancer.Theranostics. 2021 Jul 13;11(16):8112-8128. doi: 10.7150/thno.54961. eCollection 2021. Theranostics. 2021. PMID: 34335983 Free PMC article.
-
Identified Candidate Genes of Semen Trait in Three Pig Breeds Through Weighted GWAS and Multi-Tissue Transcriptome Analysis.Animals (Basel). 2025 Feb 5;15(3):438. doi: 10.3390/ani15030438. Animals (Basel). 2025. PMID: 39943208 Free PMC article.
-
Bi-allelic variants in human TCTE1/DRC5 cause asthenospermia and male infertility.Eur J Hum Genet. 2022 Jun;30(6):721-729. doi: 10.1038/s41431-022-01095-w. Epub 2022 Apr 7. Eur J Hum Genet. 2022. PMID: 35388187 Free PMC article.
-
Comparative analysis of mRNA transcripts of HT-29 cell line expressed in identical quantities for pathogenic E. coli strains UM146 and UM147 with control Escherichia coli Nissle 1917.J Genomics. 2022 Jan 1;10:1-7. doi: 10.7150/jgen.67277. eCollection 2022. J Genomics. 2022. PMID: 34976225 Free PMC article.
References
-
- Armbrust EV, Berges JA, Bowler C, Green BR, Martinez D, Putnam NH, Zhou S, Allen AE, Apt KE, Bechner M, et al. The genome of the diatom Thalassiosira pseudonana: ecology, evolution, and metabolism. Science. 2004;306:79–86. - PubMed
-
- Austin-Tse C, Halbritter J, Zariwala MA, Gilberti RM, Gee HY, Hellman N, Pathak N, Liu Y, Panizzi JR, Patel-King RS, et al. Zebrafish ciliopathy screen plus human mutational analysis identifies C21orf59 and CCDC65 defects as causing primary ciliary dyskinesia. Am J Hum Genet. 2013;93:672–686. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
