

The mechanism of variability in transcription start site selection
- PMID: 29168694
- PMCID: PMC5730371
- DOI: 10.7554/eLife.32038
The mechanism of variability in transcription start site selection
Abstract
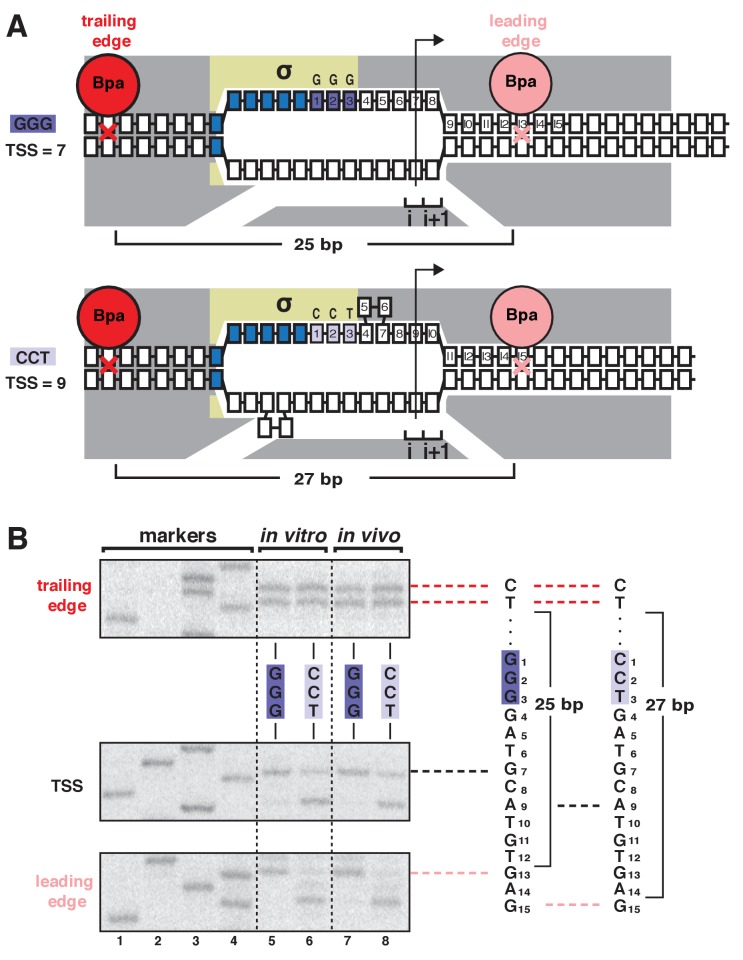
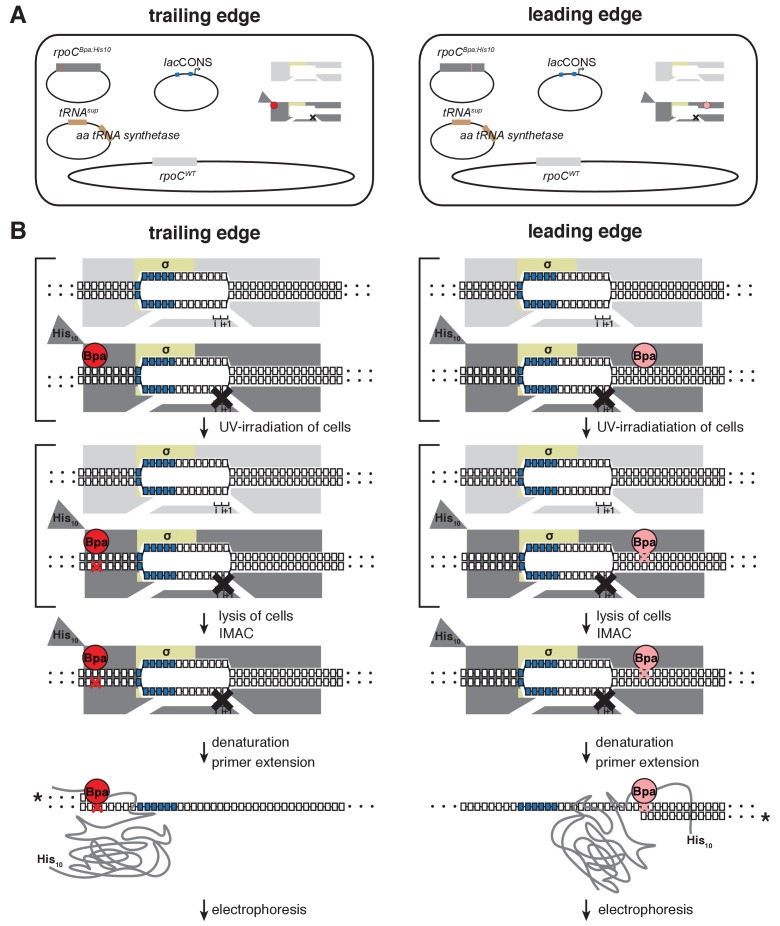
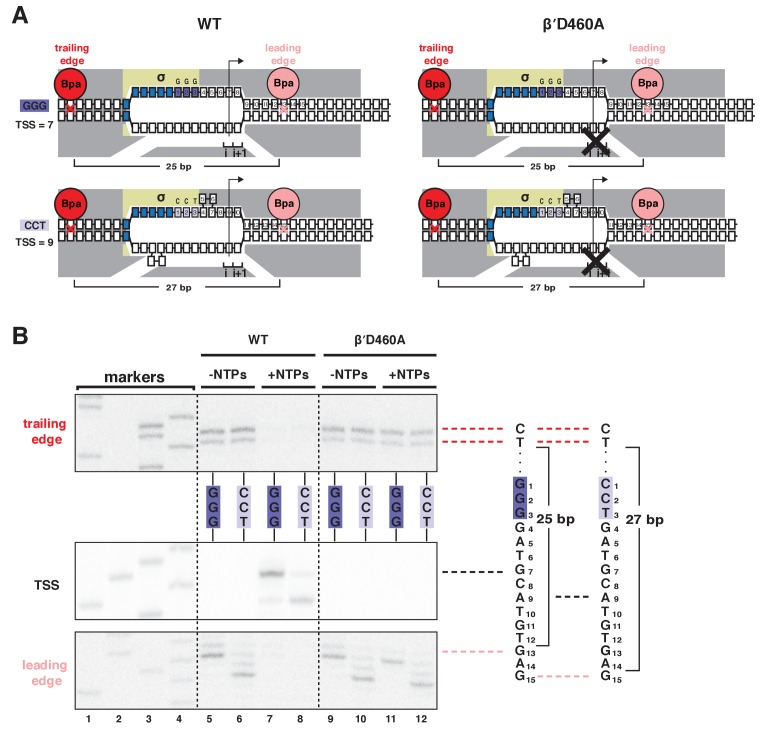
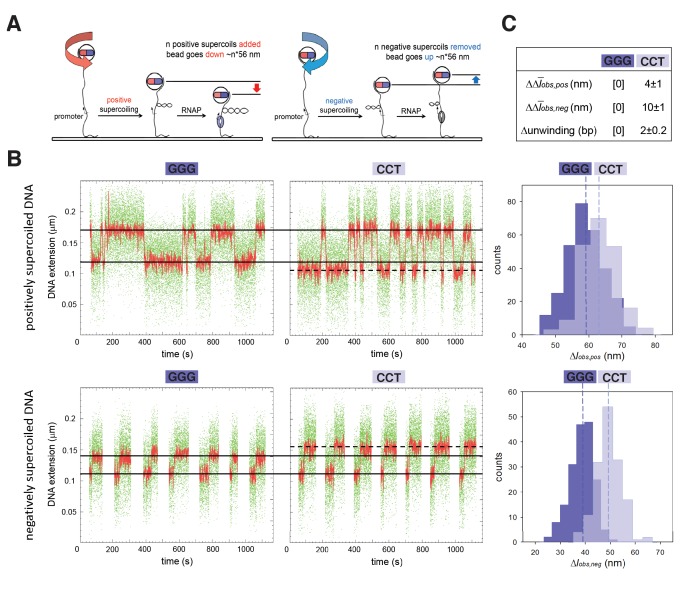
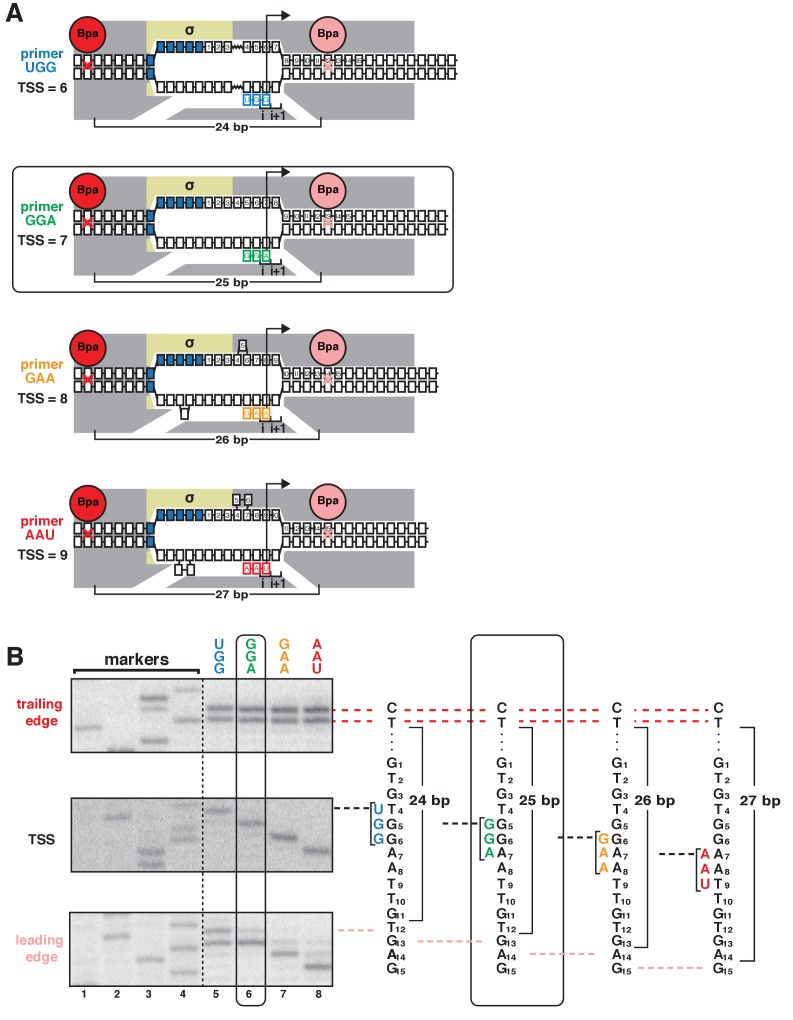
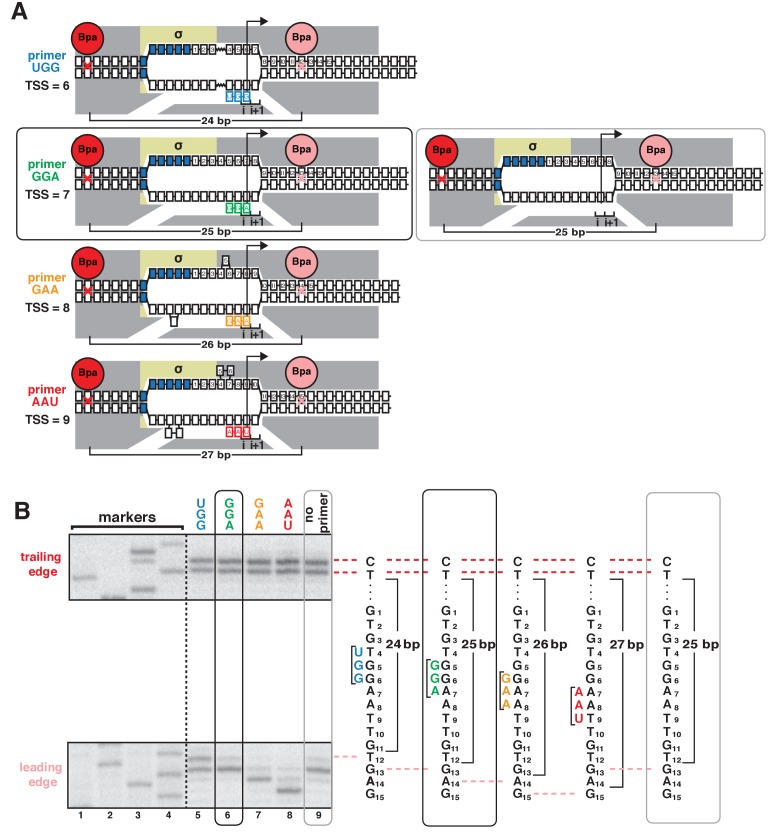
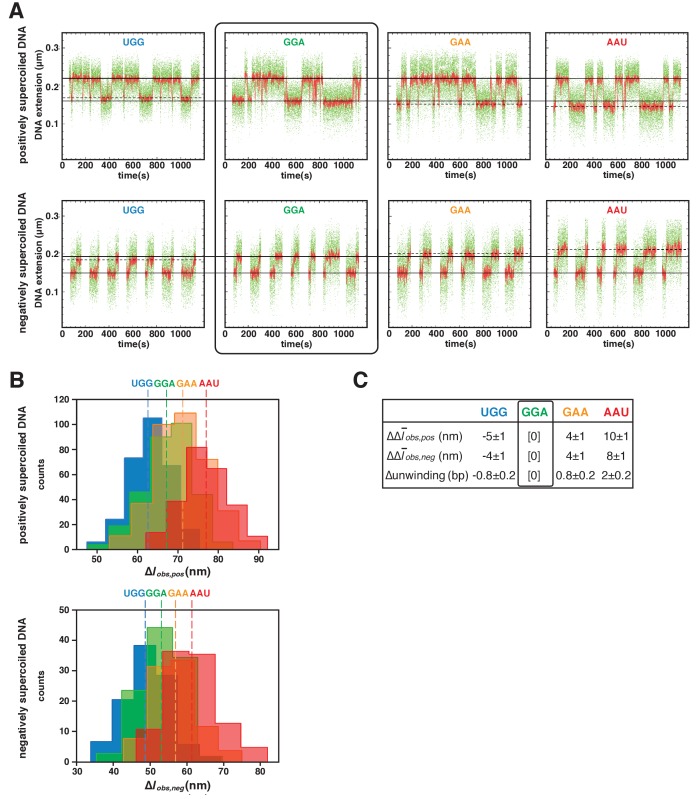
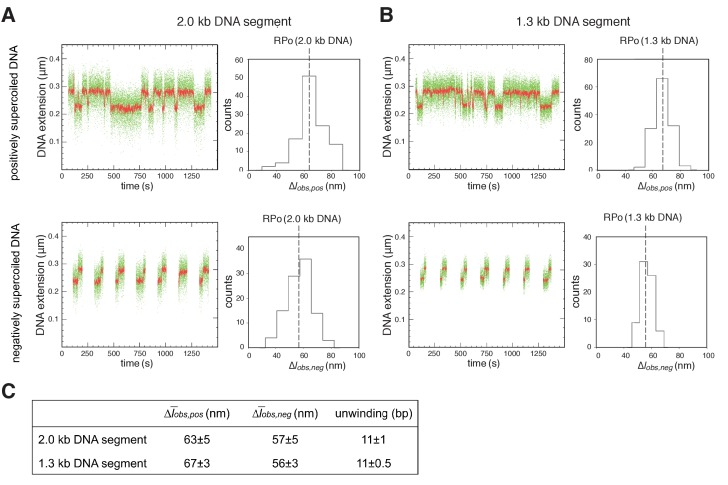
During transcription initiation, RNA polymerase (RNAP) binds to promoter DNA, unwinds promoter DNA to form an RNAP-promoter open complex (RPo) containing a single-stranded 'transcription bubble,' and selects a transcription start site (TSS). TSS selection occurs at different positions within the promoter region, depending on promoter sequence and initiating-substrate concentration. Variability in TSS selection has been proposed to involve DNA 'scrunching' and 'anti-scrunching,' the hallmarks of which are: (i) forward and reverse movement of the RNAP leading edge, but not trailing edge, relative to DNA, and (ii) expansion and contraction of the transcription bubble. Here, using in vitro and in vivo protein-DNA photocrosslinking and single-molecule nanomanipulation, we show bacterial TSS selection exhibits both hallmarks of scrunching and anti-scrunching, and we define energetics of scrunching and anti-scrunching. The results establish the mechanism of TSS selection by bacterial RNAP and suggest a general mechanism for TSS selection by bacterial, archaeal, and eukaryotic RNAP.
Keywords: E. coli; RNA polymerase; biophysics; chromosomes; genes; magnetic tweezers; p-benzoyl-L-phenylalanine (Bpa); protein-DNA photocrosslinking; scrunching; single-molecule biophysics; structural biology.
Conflict of interest statement
No competing interests declared.
Figures
Similar articles
-
Interactions between RNA polymerase and the core recognition element are a determinant of transcription start site selection.Proc Natl Acad Sci U S A. 2016 May 24;113(21):E2899-905. doi: 10.1073/pnas.1603271113. Epub 2016 May 9. Proc Natl Acad Sci U S A. 2016. PMID: 27162333 Free PMC article.
-
Multiplexed protein-DNA cross-linking: Scrunching in transcription start site selection.Science. 2016 Mar 4;351(6277):1090-3. doi: 10.1126/science.aad6881. Science. 2016. PMID: 26941320 Free PMC article.
-
Open complex scrunching before nucleotide addition accounts for the unusual transcription start site of E. coli ribosomal RNA promoters.Proc Natl Acad Sci U S A. 2016 Mar 29;113(13):E1787-95. doi: 10.1073/pnas.1522159113. Epub 2016 Mar 14. Proc Natl Acad Sci U S A. 2016. PMID: 26976590 Free PMC article.
-
Open complex DNA scrunching: A key to transcription start site selection and promoter escape.Bioessays. 2017 Feb;39(2):10.1002/bies.201600193. doi: 10.1002/bies.201600193. Epub 2017 Jan 4. Bioessays. 2017. PMID: 28052345 Free PMC article. Review.
-
How to switch the motor on: RNA polymerase initiation steps at the single-molecule level.Protein Sci. 2017 Jul;26(7):1303-1313. doi: 10.1002/pro.3183. Epub 2017 May 12. Protein Sci. 2017. PMID: 28470684 Free PMC article. Review.
Cited by
-
Step-by-Step Regulation of Productive and Abortive Transcription Initiation by Pyrophosphorolysis.J Mol Biol. 2022 Jul 15;434(13):167621. doi: 10.1016/j.jmb.2022.167621. Epub 2022 May 6. J Mol Biol. 2022. PMID: 35533764 Free PMC article.
-
XACT-seq: A photocrosslinking-based technique for detection of the RNA polymerase active-center position relative to DNA in Escherichia coli.STAR Protoc. 2021 Oct 8;2(4):100858. doi: 10.1016/j.xpro.2021.100858. eCollection 2021 Dec 17. STAR Protoc. 2021. PMID: 34693360 Free PMC article.
-
Structural and mechanistic basis of reiterative transcription initiation.Proc Natl Acad Sci U S A. 2022 Feb 1;119(5):e2115746119. doi: 10.1073/pnas.2115746119. Proc Natl Acad Sci U S A. 2022. PMID: 35082149 Free PMC article.
-
Transient-state kinetic analysis of multi-nucleotide addition catalyzed by RNA polymerase I.Biophys J. 2021 Oct 19;120(20):4378-4390. doi: 10.1016/j.bpj.2021.09.008. Epub 2021 Sep 10. Biophys J. 2021. PMID: 34509510 Free PMC article.
-
The Context-Dependent Influence of Promoter Sequence Motifs on Transcription Initiation Kinetics and Regulation.J Bacteriol. 2021 Mar 23;203(8):e00512-20. doi: 10.1128/JB.00512-20. Print 2021 Mar 23. J Bacteriol. 2021. PMID: 33139481 Free PMC article. Review.
References
-
- Artsimovitch I, Svetlov V, Murakami KS, Landick R. Co-overexpression of Escherichia coli RNA polymerase subunits allows isolation and analysis of mutant enzymes lacking lineage-specific sequence insertions. Journal of Biological Chemistry. 2003;278:12344–12355. doi: 10.1074/jbc.M211214200. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
