

R-Ras-Akt axis induces endothelial lumenogenesis and regulates the patency of regenerating vasculature
- PMID: 29170374
- PMCID: PMC5700916
- DOI: 10.1038/s41467-017-01865-x
R-Ras-Akt axis induces endothelial lumenogenesis and regulates the patency of regenerating vasculature
Abstract
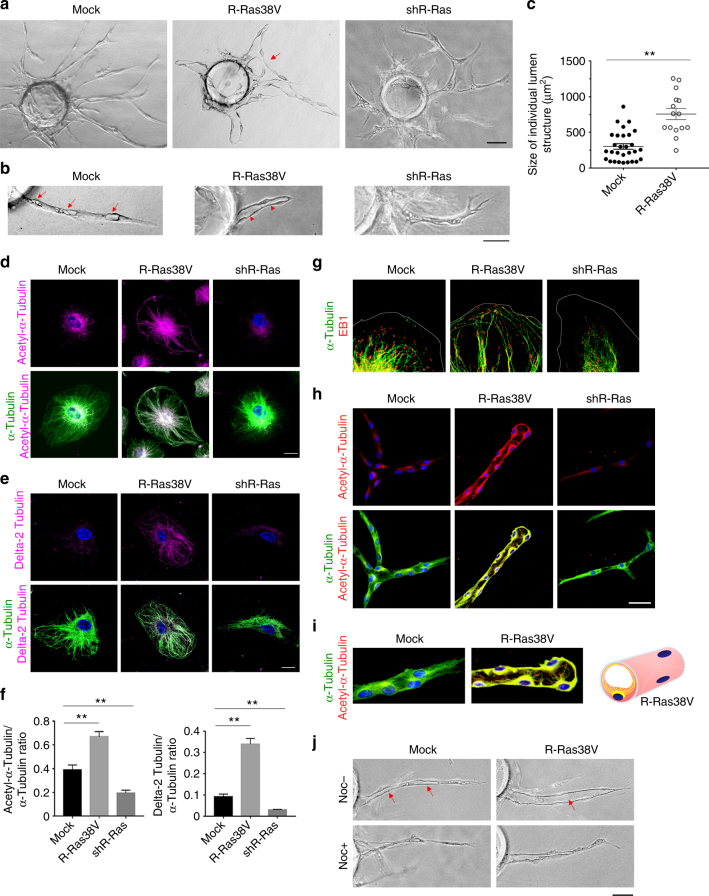
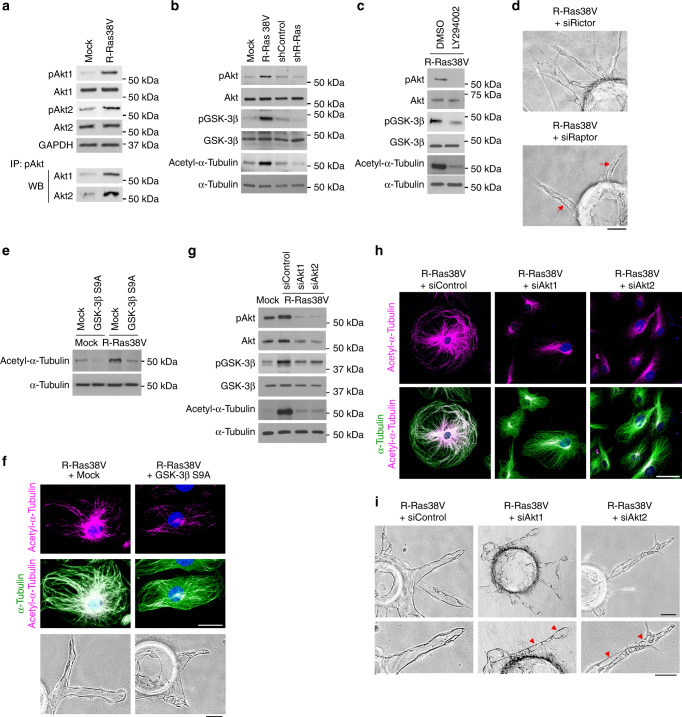
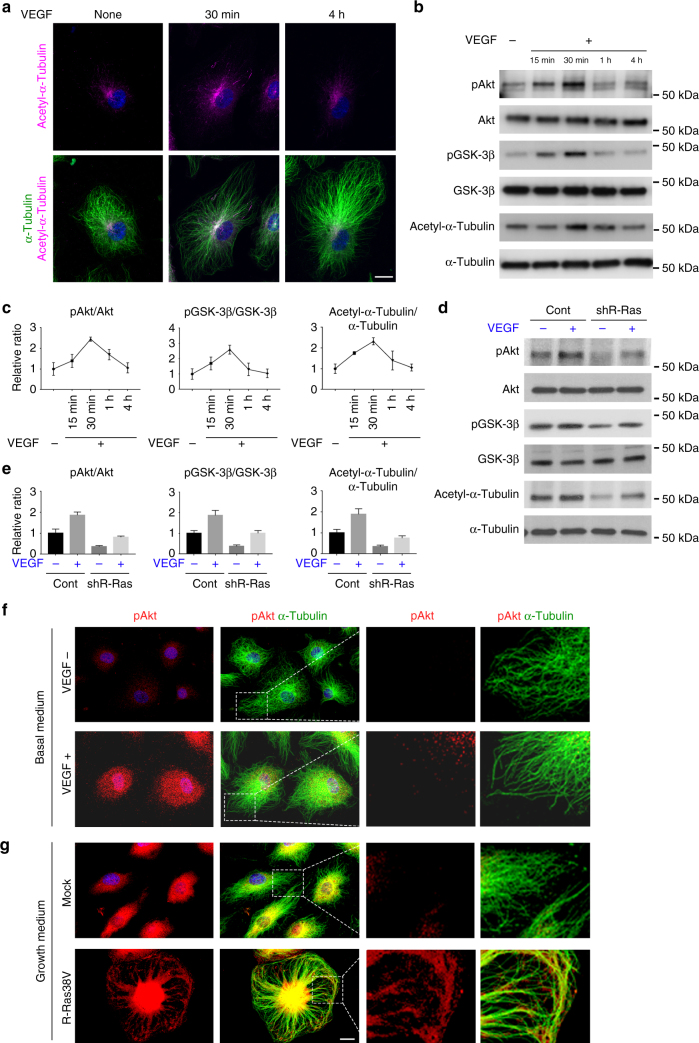
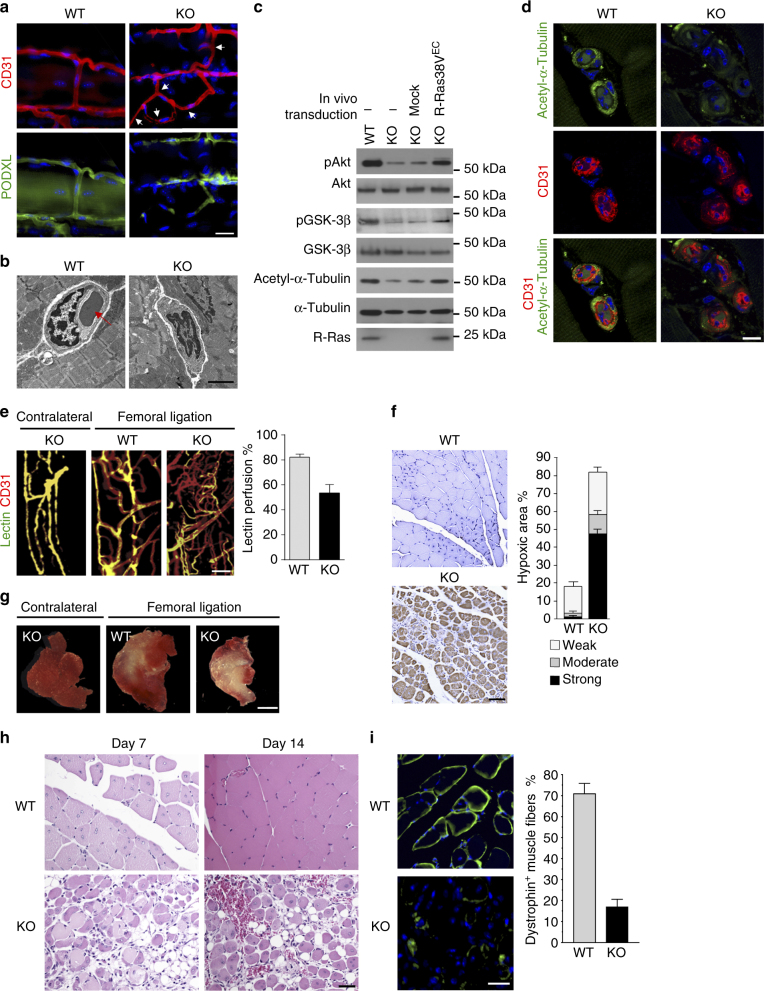
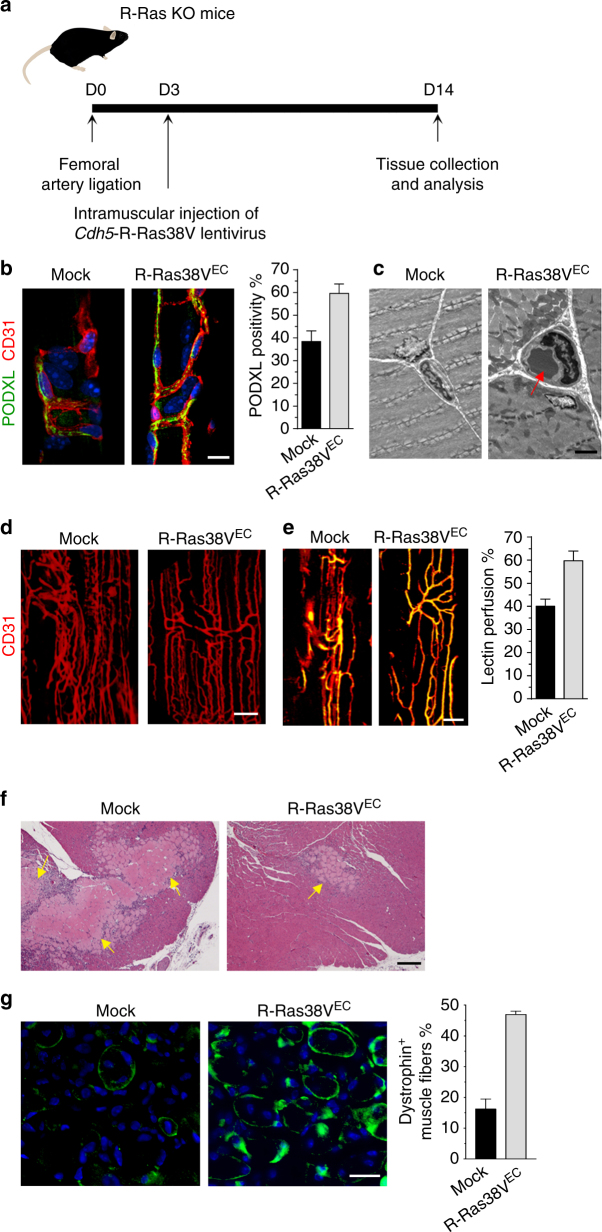
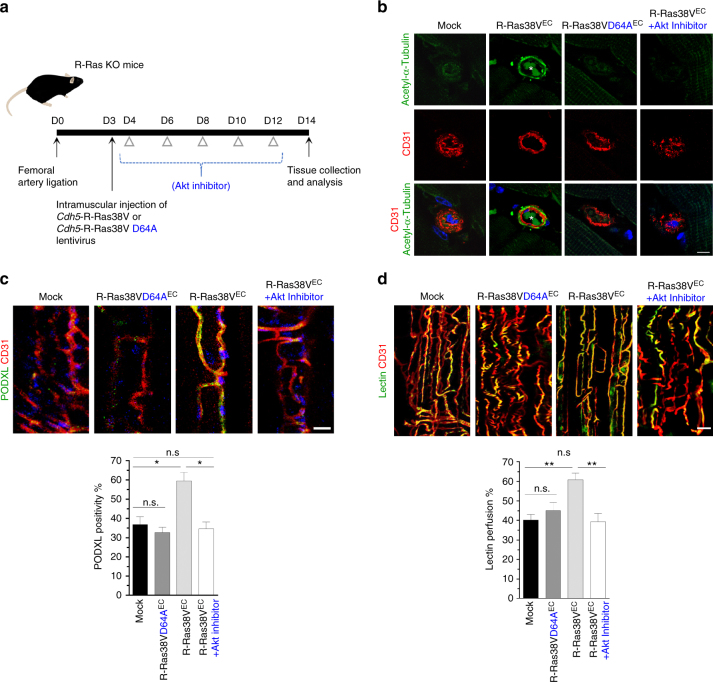
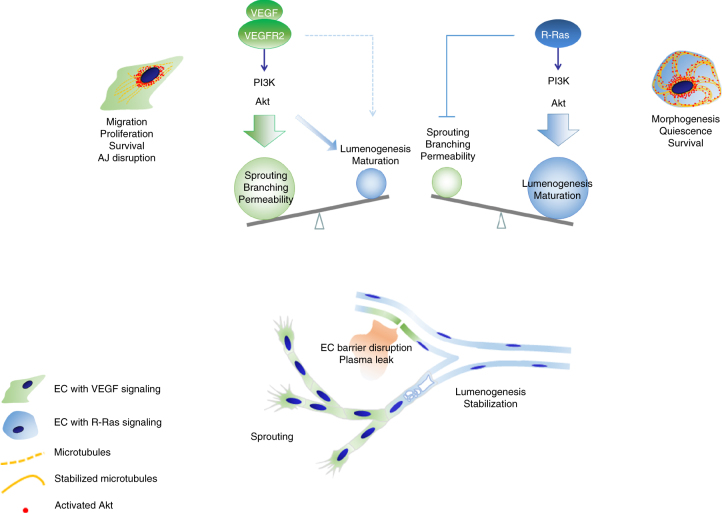
The formation of endothelial lumen is fundamental to angiogenesis and essential to the oxygenation of hypoxic tissues. The molecular mechanism underlying this important process remains obscure. Here, we show that Akt activation by a Ras homolog, R-Ras, stabilizes the microtubule cytoskeleton in endothelial cells leading to endothelial lumenogenesis. The activation of Akt by the potent angiogenic factor VEGF-A does not strongly stabilize microtubules or sufficiently promote lumen formation, hence demonstrating a distinct role for the R-Ras-Akt axis. We show in mice that this pathway is important for the lumenization of new capillaries and microvessels developing in ischemic muscles to allow sufficient tissue reperfusion after ischemic injury. Our work identifies a role for Akt in lumenogenesis and the significance of the R-Ras-Akt signaling for the patency of regenerating blood vessels.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
