

Heme enables proper positioning of Drosha and DGCR8 on primary microRNAs
- PMID: 29170488
- PMCID: PMC5700927
- DOI: 10.1038/s41467-017-01713-y
Heme enables proper positioning of Drosha and DGCR8 on primary microRNAs
Erratum in
-
Publisher Correction: Heme enables proper positioning of Drosha and DGCR8 on primary microRNAs.Nat Commun. 2018 Sep 18;9(1):3852. doi: 10.1038/s41467-018-06426-4. Nat Commun. 2018. PMID: 30228298 Free PMC article.
Abstract
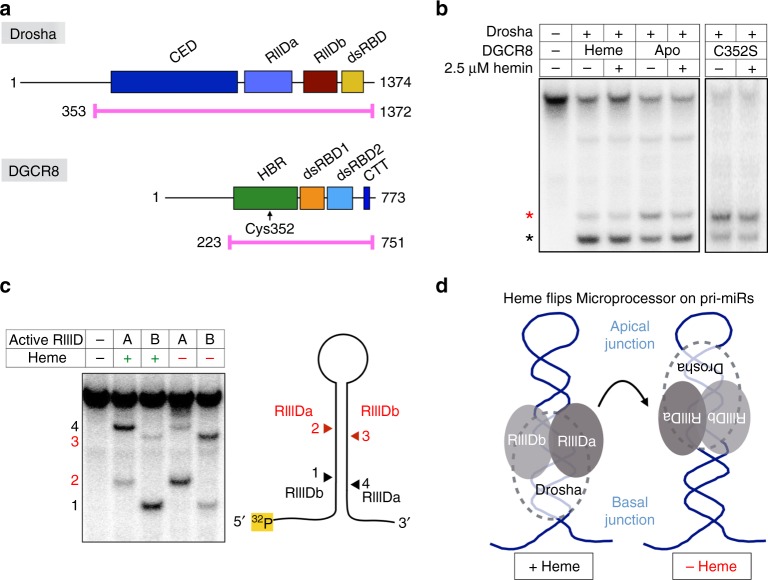
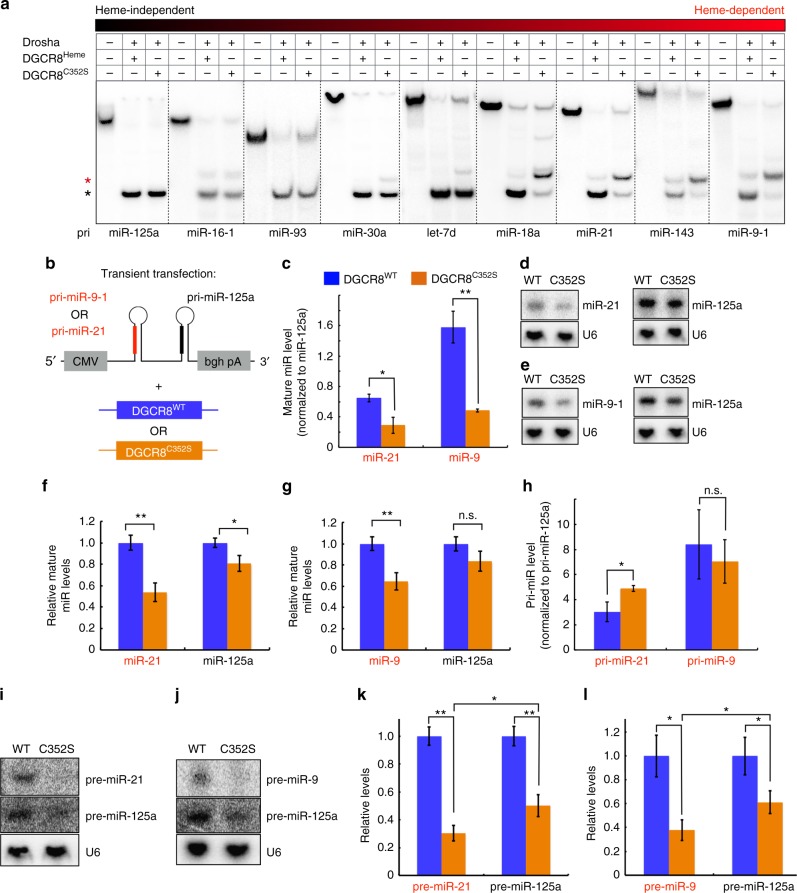
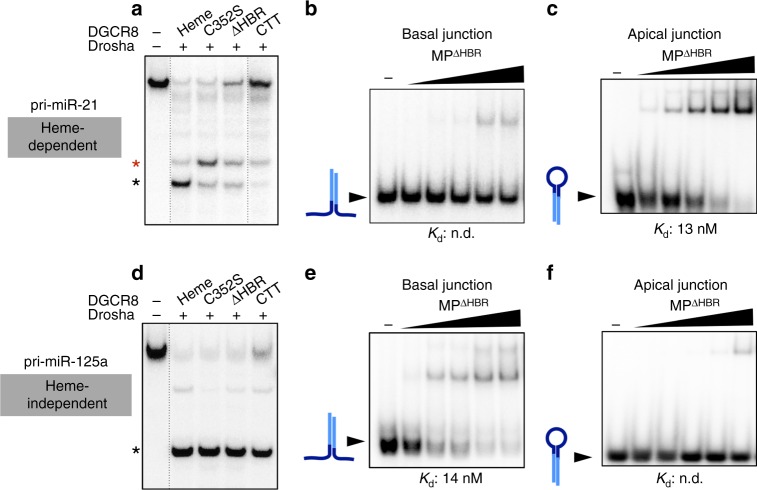
MicroRNAs regulate the expression of many proteins and require specific maturation steps. Primary microRNA transcripts (pri-miRs) are cleaved by Microprocessor, a complex containing the RNase Drosha and its partner protein, DGCR8. Although DGCR8 is known to bind heme, the molecular role of heme in pri-miR processing is unknown. Here we show that heme is critical for Microprocessor to process pri-miRs with high fidelity. Furthermore, the degree of inherent heme dependence varies for different pri-miRs. Heme-dependent pri-miRs fail to properly recruit Drosha, but heme-bound DGCR8 can correct erroneous binding events. Rather than changing the oligomerization state, heme induces a conformational change in DGCR8. Finally, we demonstrate that heme activates DGCR8 to recognize pri-miRs by specifically binding the terminal loop near the 3' single-stranded segment.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
