

Review
doi: 10.1016/j.preteyeres.2017.11.002.
Epub 2017 Nov 22.
Retinal energy demands control vascular supply of the retina in development and disease: The role of neuronal lipid and glucose metabolism
Affiliations
- PMID: 29175509
- PMCID: PMC5963988
- DOI: 10.1016/j.preteyeres.2017.11.002
Item in Clipboard
Review
Retinal energy demands control vascular supply of the retina in development and disease: The role of neuronal lipid and glucose metabolism
Prog Retin Eye Res.
2018 May.
No abstract available
Figures

(a) Similar radial distribution of neurons (RGCs marked with leptin; blue), glia (astrocytes labeled with GFAP; green) and blood vessels (Dextran; red). Arrow points to the optic nerve. (b) Migratory tip cell, which does not proliferate (star; nucleus) with adjacent proliferating stalk cell in cellular division (phospho-histone-H3, green with arrow). (c) Astrocytes lay down a path for growing vessels, starting before birth (E20, left) and completed at birth (P0); vessels are red (right). (d) Inner retinal vascular development begins with the formation of the superficial vascular plexus (1 and e). Formation of the deep vascular plexus (2) corresponds temporally to the maturation of photoreceptors (f). The intermediate vascular plexus forms last (3), likely from vessels of the superficial vascular plexus. (f) Timeline of photoreceptor development from the terminal mitosis of rod precursors to the onset of rhodopsin expression. Mice open their eyes between P12-15, and the outer plexiform layer has developed by P14. Although some rod activity is detectable at P12, photoreceptor outer segments continue to mature until P19-25. Figure modified, with permission, from (Dorrell and Friedlander, 2006; Gariano and Gardner, 2005; Gerhardt et al., 2003; Morrow et al., 1998).

(a,b) Distribution of cones (c) and rods (r) in the human retina, relative to the optic nerve. Cone density increases in the macula and peaks in the fovea. (c) Macaque retina stained for cytochrome c oxidase (COX) or Na + K + ATPase, markers of mitochondria or areas of higher energy consumption in the retina. Inner segments (IS of photoreceptors strongly express both markers. NFL: Nerve fiber layer, GCL: ganglion cell layer, IPL: Inner plexiform layer, INL: Inner nuclear layer, OPL: Outer plexiform layer, ONL: Outer nuclear layer, IS: inner segment, OS: outer segment. (d) Cone inner segments (CIS) contain much larger and more densely packed mitochondria than rod photoreceptor, by electron microscopy. Figure modified, with permission, from (Curcio et al., 1990; Kageyama and Wong-Riley, 1984; Wong-Riley et al., 1998).
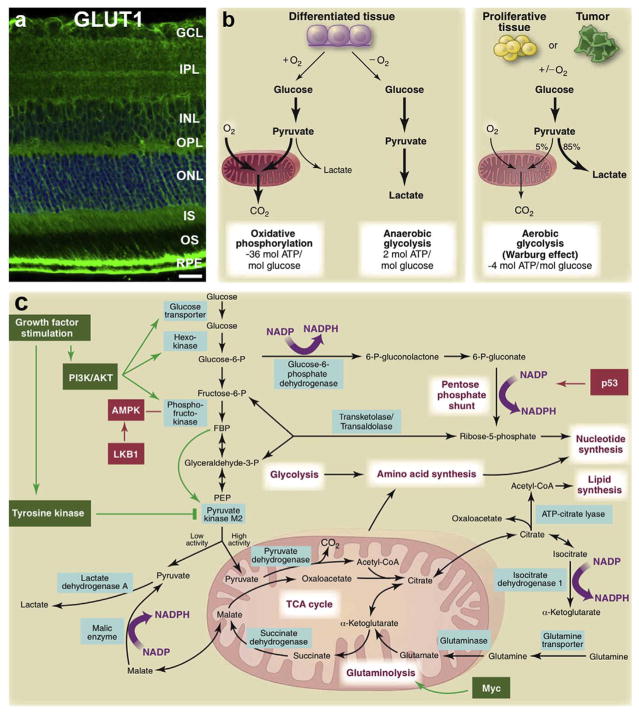
(a) Distribution of GLUT1, the main glucose transporter of the retina. (b) Schematic representation of oxidative phosphorylation, anaerobic glycolysis, and aerobic glycolysis (Warburg effect). In the presence of oxygen, differentiated tissues metabolize glucose to pyruvate via glycolysis. Pyruvate is then metabolized in mitochondria by oxidative phosphorylation. The electron transport chain requires oxygen to completely oxidize glucose. When oxygen is scarce, cells redirect pyruvate away from mitochondria to produce lactate (anaerobic glycolysis). Anaerobic glycolysis allows glycolysis to continue by cycling NADH back to NAD+but limits ATP production compared to oxidative phosphorylation. Warburg observed that cancer cells convert most glucose to lactate, irrespective of the presence of oxygen (aerobic glycolysis). The ‘Warburg effect’ is also observed in non cancerous tissues, such as the retina, which, though non proliferative have continuous replacement of outer segments and behave metabolically as “proliferative” cells. Aerobic glycolysis which is less energy efficient may provide building blocks required for growth. In proliferating cells, glucose is in part diverted to biosynthetic pathways upstream of pyruvate production. (c) This schematic summarizes glucose metabolism including glycolysis, oxidative phosphorylation, the pentose phosphate pathway, and glutamine metabolism in “proliferating” cells. Growth factor signaling, such as VEGF, leads to both tyrosine kinase and AKT/PI3K activation. In doing so, growth factors promote glucose uptake and flux through the early part of glycolysis, while inhibiting the late steps; they force glycolytic intermediates towards biosynthetic pathways of macromolecules essential for cell “proliferation” and NADPH production. Metabolic pathways are labeled in purple, and the enzymes controlling critical metabolic steps are shown in blue boxes. Figure modified, with permission, from (Gospe et al., 2010; Vander Heiden et al., 2009).
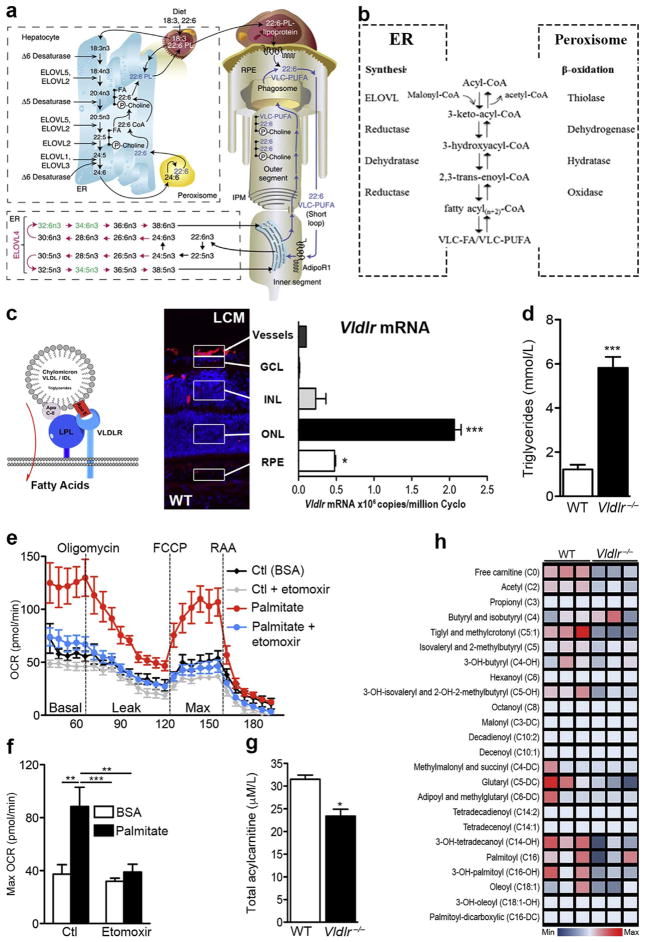
(a) Schematic of the formation of VLC-PUFAs. The elongation steps catalyzed by ELOVL4 are unique to photoreceptor inner segments (in red). Shed photoreceptor apical disks are phagocytosed by RPE. DHA (22:6) and VLD-PUFAs are recycled back to photoreceptor inner segments. C, carbons; ELOVL, elongase of the very long-chain fatty acids; ER, endoplasmic reticulum; IPM, interphotoreceptor matrix; PL, phospholipid; RPE, retinal pigment epithelium. (b) Fatty acid elongation and β-oxidation pathways. Each round of elongation involves four successive steps in the endoplasmic reticulum. Elongated fatty acyl-CoA product may undergo further rounds of elongation, be released for use in the cell, or, β-oxidized in the peroxisome. (c) VLDL receptors bind triglyceride-rich chylomicrons and VLDLs that express Apo-E, allowing lipoprotein lipase (LPL) to release long-chain fatty acids. VLDLR is highly expressed in photoreceptors (outer nuclear layer; ONL) by laser capture microdissection (LCM and qRT-PCR). GCL: ganglion cell layer, INL: inner nuclear layer, RPE: retinal pigment epithelium. n = 3 retinas, scale: 50 μm. (d) Circulating plasma triglyceride levels in WT and Vldlr−/− mice. P < 0.0001. (e) Oxygen consumption rate (OCR) and (f) maximal OCR of wild-type (WT) retinas provided with long-chain fatty acid (FA) palmitate or control (Ctl: bovine serum albumin or BSA) in the presence or absence of FA oxidation inhibitor, etomoxir (40 μM). n=6–8 retinas. (g) Total acylcarnitine levels (P = 0.0108) and (h) metabolite array of FA β-oxidation, measured by LC/MS/MS. n = 3 animal retinas. Figure modified, with permission, from (Joyal et al., 2016; Rice et al., 2015).
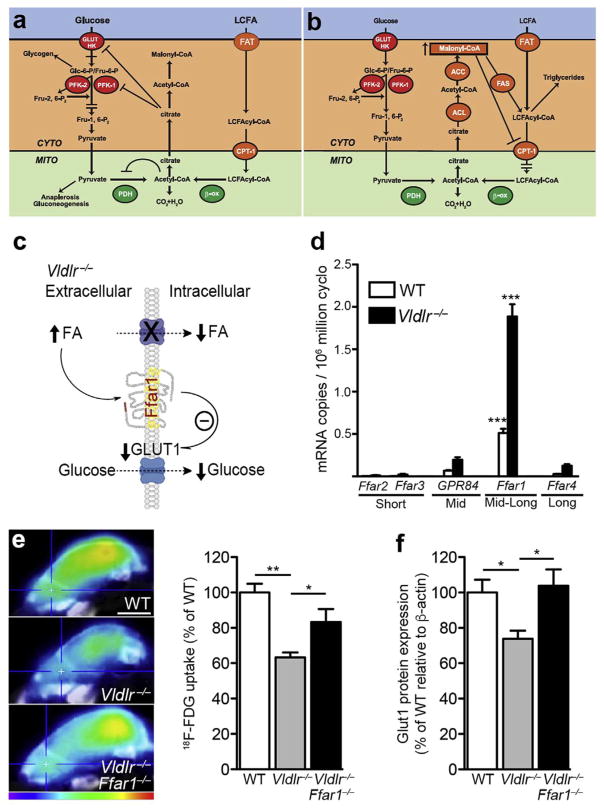
(a) Randle cycle: inhibition of glucose utilization by fatty acid oxidation. Accumulation of acetyl-CoA and NADH from FA oxidation inhibits pyruvate dehydrogenase (PDH), whereas cytosolic citrate regulates 6-phosphofructo-1-kinase (PFK) activity. Glucose uptake regulation is not fully explained by the Randle cycle. (b) Randle cycle: inhibition of fatty acid oxidation by glucose. Malonyl-CoA, which is produced by ACC when glucose is abundant, governs the expression of CPT1, hence regulating the entry of long-chain FA into the mitochondria. This effect re-routes fatty acids toward esterification and storage. CYTO: cytosol; MITO: mitochondria; GLUT: glucose transporter; HK: hexokinase; Glc-6-P: glucose 6-phosphate; Fru-6-P: fructose 6-phosphate; CPTI: carnitine palmitoyltransferase I; β-ox: β-oxidation, ACC: Acetyl-CoA carboxylase, ACL: ATP-citrate lyase; FAS, fatty acid synthase. (c) Elevated circulating fatty acid levels, as seen in Vldlr−/− retina, activate fatty acid receptors (such as Ffar1) that suppress GLUT1 expression and glucose uptake when lipids are abundant. (d) FA sensing receptors are expressed in WT and Vldlr−/−retinas (qRT-PCR). ONL: outer nuclear layer, INL: inner nuclear layer, GCL: ganglion cell layer. n = 3 animal retinas. (g) Glucose uptake (18F-FDG, scale: 4 mm) and Glut1 protein expression of WT and Vldlr−/− mice compared to littermate Vldlr−/−/Ffar1−/− mice (P16). Figure modified, with permission, from (Hue and Taegtmeyer, 2009; Joyal et al., 2016).
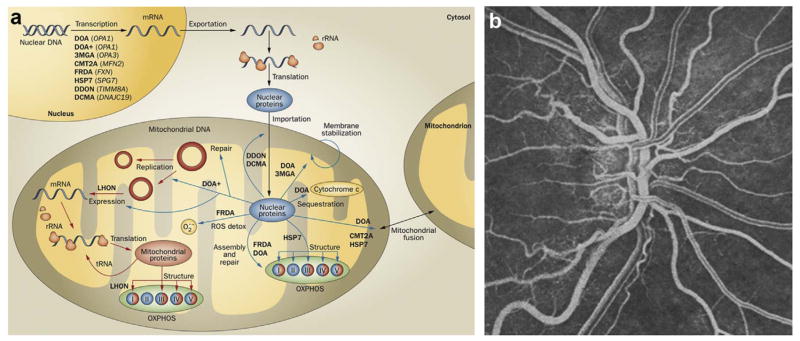
(a) Cellular homeostasis is under the dual control of nuclear (blue) and mitochondrial genomes. Mitochondrial diseases (bold text), many of which impact vision, arise from mutations in genes (italic text) from either genome. Mitochondrial DNA encodes 13 structural subunits of the electron transport chain (complexes I, III, IV, and V) and the RNA needed for gene translation. All other mitochondrial proteins are encoded by the nuclear genome. Abbreviations: CMT2A, Charcot-Marie-Tooth disease, type 2A; DCMA, dilated cardiomyopathy with ataxia; DDON, deafness, dystonia and optic neuropathy; DOA, dominant optic atrophy; DOA+, dominant optic atrophy-plus syndrome; FRDA, Friedreich ataxia; HSP7, hereditary spastic paraplegia, type 7; LHON, Leber hereditary optic neuropathy; OXPHOS, oxidative phosphorylation; ROS detox, reactive oxygen species detoxification; rRNA, ribosomal RNA; tRNA, transfer RNA; 3MGA, 3-methylglutaconic aciduria, type III. (b) Fluorescein angiography of fundus during the acute stage of LHON, showing tortuous vessels and telangiectasias. Energy metabolism has a direct impact on the retinal vascular phenotype. Figure modified, with permission, from (Newman, 2012).

(a) Dual shortage of glucose (b, metabolized to pyruvate) and FA uptake reduces acetyl-CoA (c, estimated by measuring acetylcarnitine) and (d) TCA (Krebs) cycle intermediate α-KG in Vldlr−/− retina (LC/MS/MS). Together with oxygen (O2), α-KG is an essential co-activator of propyl-hydroxylase dehydrogenase (PHD) that tags HIF-1α for degradation by proline hydroxylation (hydroxyproline). (e) Levels of hydroxyproline residues in WT and Vldlr−/− retinas measured by LC/MS/MS (n = 15 WT, 12 Vldlr−/− animal retinas, P = 0.0004). (f) Hif1a retinal expression in Vldlr−/− photoreceptor layer (P12 retinal flat mounts, Scale: 100 μm; left: extended focus; middle and right panels: 3D confocal IHC, n = 3) where (g) Vegfa was then also secreted and localized (P16 retinal flat mounts, Scale: 100 μm; left: extended focus; middle and right panels: 3D confocal IHC, n = 3 retinas). (h) Human subjects with AMD, either retinal angiomatous proliferation (RAP, n =3) or choroidal neovascularization (CNV, n = 7) had higher VEGFA vitreous levels by ELISA compared to control subjects without pathologic neovessels (macular hole; n = 8). Results are presented as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Figure modified, with permission, from (Joyal et al., 2016).
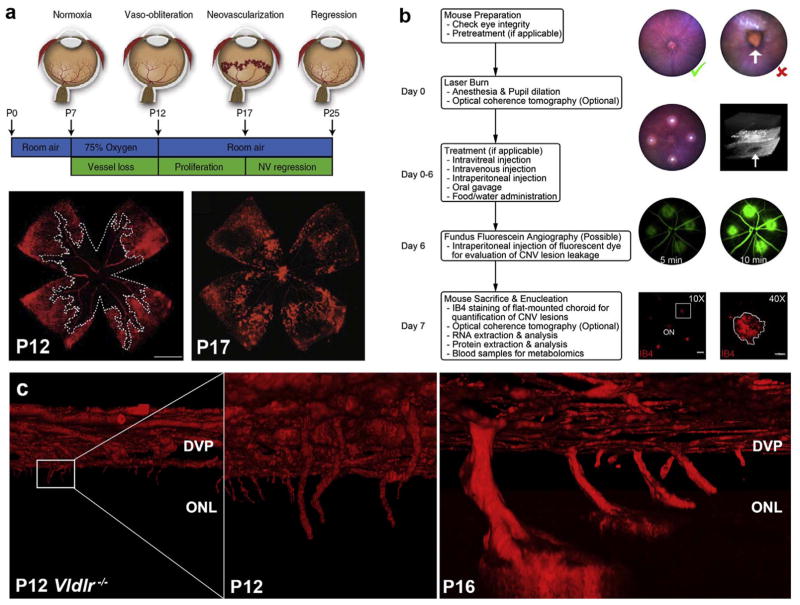
(a) Oxygen-induced retinopathy: the mouse model of proliferative retinopathy. Neonatal mice are exposed to 75% oxygen from P7 to P12, which induces loss of immature retinal vessels, leading to a central zone of vaso-obliteration (VO). Mice are returned to room air at P12, and the central avascular retina becomes hypoxic, triggering both vascular regrowth and the formation of pathologic neovascular (NV) tufts culminating at P17. (b) Laser-induced choroidal neovascularization (CNV) model. Detailed flowchart of the procedure to obtain consistent laser-induced CNV lesions. Flat-mounted choroid vessels are stained with IB4 at day 7 after laser photocoagulation. 10× Scale bar: 200 μm, 40× Scale bar: 50 μm. ON, optic nerve. (c) Mice model of retinal angiomatous proliferation (RAP): Vldlr−/− mice. Mice deficient for Vldlr spontaneously develop pathological RAP-like vascular lesions invading the photoreceptor layer at P12 and reaching the RPE at P16. Figure modified, with permission, from (Connor et al., 2009; Gong et al., 2015; Joyal et al., 2016).
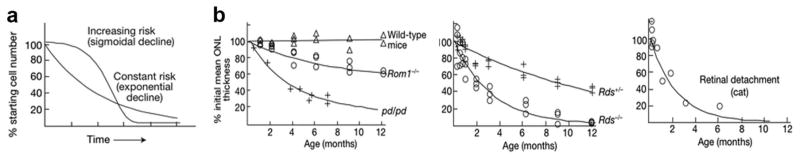
(a) Irrespective of the underlying cause, the risk of neuronal death appears constant, manifesting as an exponential decline in cell number. If the risk of neuronal death were increasing, as presumed with mitochondrial diseases and the accumulation of free radicals over time, one would observe a sigmoidal decline in cell number, which is rarely reported. Compensatory pathways must, therefore, exist to palliate for neuronal death. (b) Examples of the exponential decline of photoreceptor degeneration. Wild-type and Rom1−/− mice, and photoreceptor dysplasia (pd/pd) in miniature schnauzers (left). Retinal degeneration of heterozygous (Rds+/−) and homozygous mice (Rds−/−) carrying a null mutation in the gene encoding peripherin/rds (middle). Experimental retinal detachment in the cat (right). Figure modified, with permission, from (Clarke et al., 2000).
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
