

Dietary energy drives the dynamic response of bovine rumen viral communities
- PMID: 29179741
- PMCID: PMC5704599
- DOI: 10.1186/s40168-017-0374-3
Dietary energy drives the dynamic response of bovine rumen viral communities
Abstract
Background: Rumen microbes play a greater role in host energy acquisition than that of gut-associated microbes in monogastric animals. Although genome-enabled advancements are providing access to the vast diversity of uncultivated microbes, our understanding of variables shaping rumen microbial communities is in its infancy. Viruses have been shown to impact microbial populations through a myriad of processes, including cell lysis and reprogramming of host metabolism. However, little is known about the processes shaping the distribution of rumen viruses or how viruses may modulate microbial-driven processes in the rumen. To this end, we investigated how rumen bacterial and viral community structure and function responded in five steers fed four randomized dietary treatments in a crossover design.
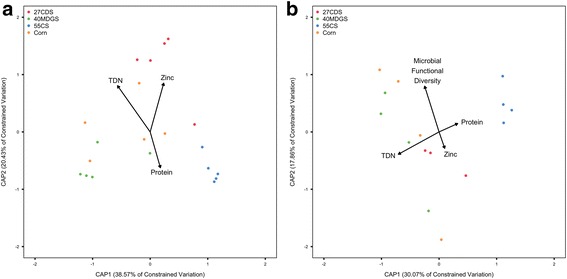
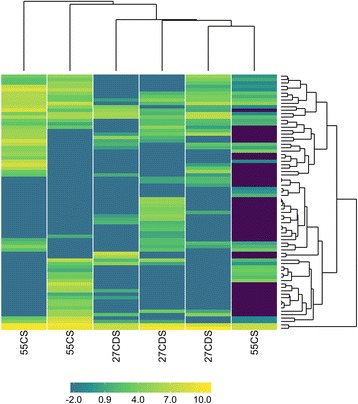
Results: Total digestible nutrients (TDN), a measure of dietary energy, best explained the variation in bacterial and viral communities. Additional ecological drivers of viral communities included dietary zinc content and microbial functional diversity. Using partial least squares regression, we demonstrate significant associations between the abundances of 267 viral populations and variables driving the variation in rumen viral communities. While rumen viruses were dynamic, 14 near ubiquitous viral populations were identified, suggesting the presence of a core rumen virome largely comprised of novel viruses. Moreover, analysis of virally encoded auxiliary metabolic genes (AMGs) indicates rumen viruses have glycosidic hydrolases to potentially augment the breakdown of complex carbohydrates to increase energy production. Other AMGs identified have a role in redirecting carbon to the pentose phosphate pathway and one carbon pools by folate to boost viral replication.
Conclusions: We demonstrate that rumen bacteria and viruses have differing responses and ecological drivers to dietary perturbation. Our results show that rumen viruses have implications for understanding the structuring of the previously identified core rumen microbiota and impacting microbial metabolism through a vast array of AMGs. AMGs in the rumen appear to have consequences for microbial metabolism that are largely in congruence with the current paradigm established in marine systems. This study provides a foundation for future hypotheses regarding the dynamics of viral-mediated processes in the rumen.
Keywords: Auxiliary metabolic genes; Phage ecology; Rumen; Viral diversity; Viral metagenome.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures





References
-
- Steinfeld H, Gerber P, Wassenaar T, Castel V, Rosales M, De Haan C. Livestock’s long shadow. Rome: FAO; 2006.
-
- Gerber PJ, Steinfeld H, Henderson B, Mottet A, Opio C, Dijkman J, et al. Tackling climate change through livestock: a global assessment of emissions and mitigation opportunities. Rome: FAO; 2013.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
