

A hereditary spastic paraplegia-associated atlastin variant exhibits defective allosteric coupling in the catalytic core
- PMID: 29180453
- PMCID: PMC5767872
- DOI: 10.1074/jbc.RA117.000380
A hereditary spastic paraplegia-associated atlastin variant exhibits defective allosteric coupling in the catalytic core
Abstract
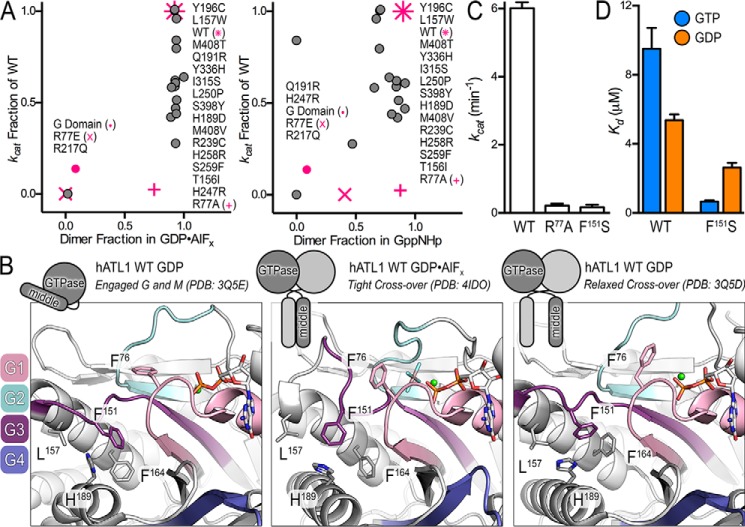
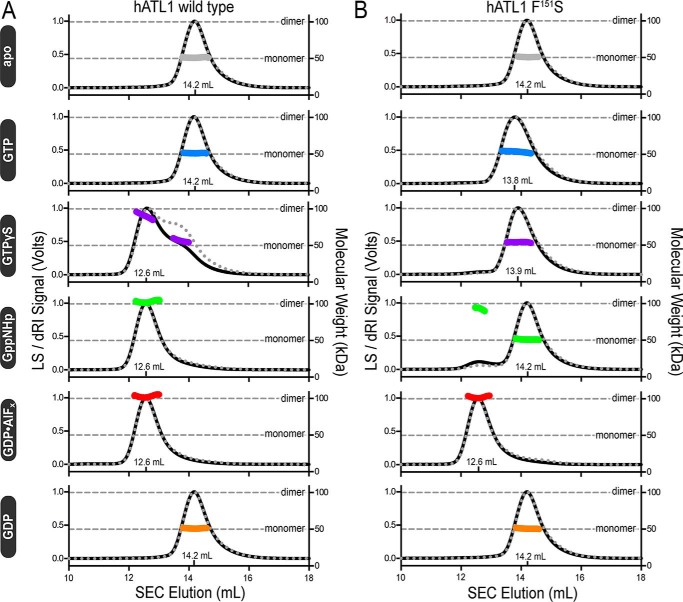
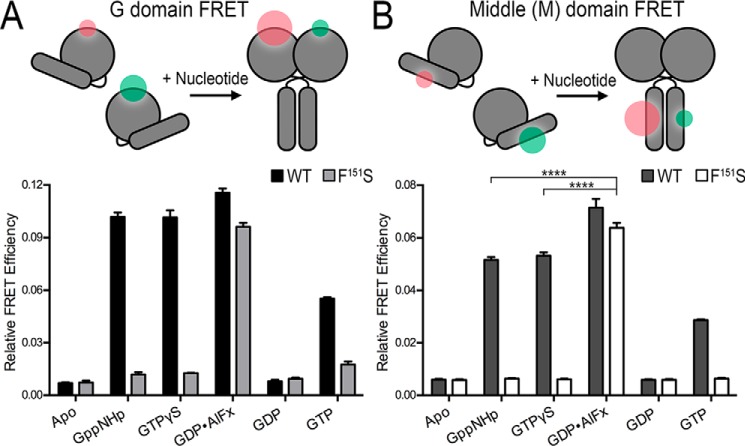
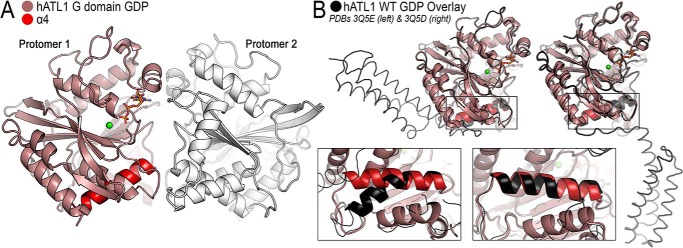
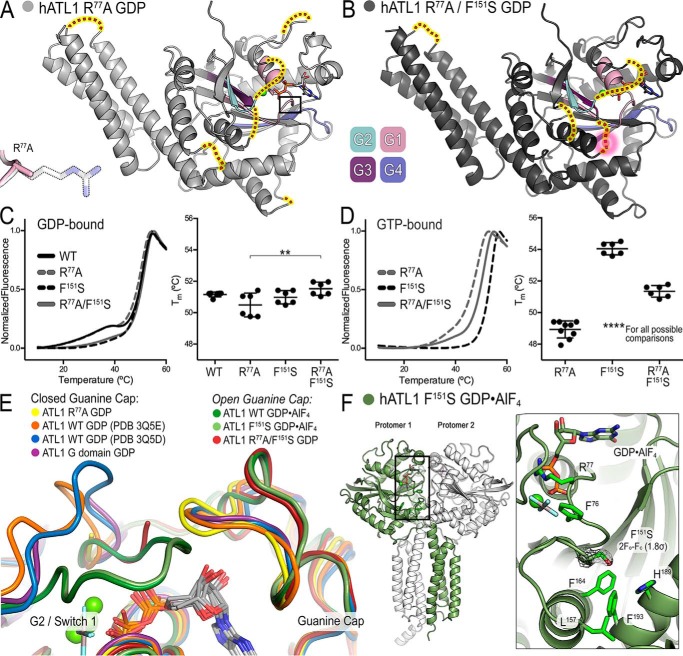
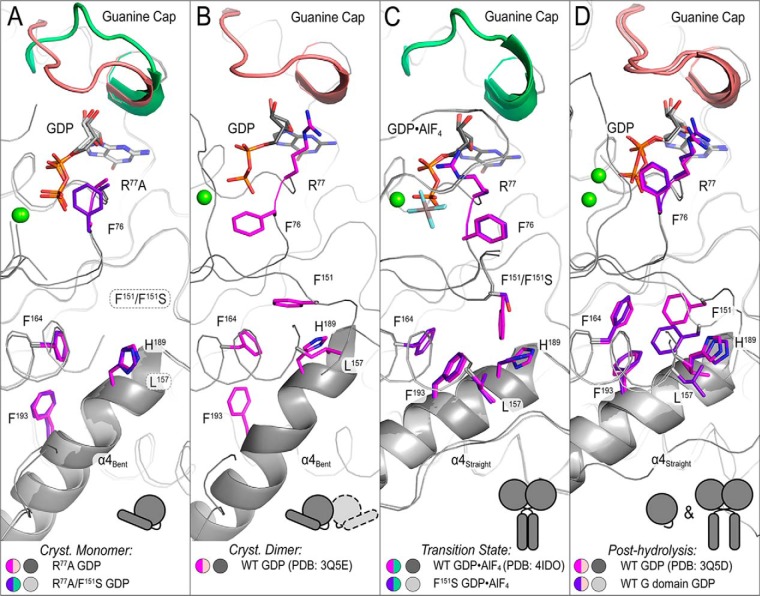
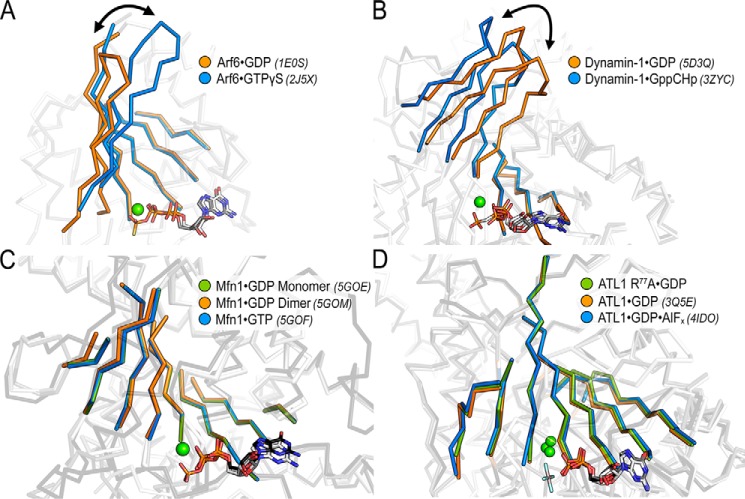
The dynamin-related GTPase atlastin (ATL) catalyzes membrane fusion of the endoplasmic reticulum and thus establishes a network of branched membrane tubules. When ATL function is compromised, the morphology of the endoplasmic reticulum deteriorates, and these defects can result in neurological disorders such as hereditary spastic paraplegia and hereditary sensory neuropathy. ATLs harness the energy of GTP hydrolysis to initiate a series of conformational changes that enable homodimerization and subsequent membrane fusion. Disease-associated amino acid substitutions cluster in regions adjacent to ATL's catalytic site, but the consequences for the GTPase's molecular mechanism are often poorly understood. Here, we elucidate structural and functional defects of an atypical hereditary spastic paraplegia mutant, ATL1-F151S, that is impaired in its nucleotide-hydrolysis cycle but can still adopt a high-affinity homodimer when bound to a transition-state analog. Crystal structures of mutant proteins yielded models of the monomeric pre- and post-hydrolysis states of ATL. Together, these findings define a mechanism for allosteric coupling in which Phe151 is the central residue in a hydrophobic interaction network connecting the active site to an interdomain interface responsible for nucleotide loading.
Keywords: GTPase; allosteric regulation; axonopathy; dynamin-related proteins; enzyme structure; hereditary spastic paraplegia; membrane fusion.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Guelly C., Zhu P.-P., Leonardis L., Papić L., Zidar J., Schabhüttl M., Strohmaier H., Weis J., Strom T. M., Baets J., Willems J., De Jonghe P., Reilly M. M., Fröhlich E., Hatz M., et al. (2011) Targeted high-throughput sequencing identifies mutations in atlastin-1 as a cause of hereditary sensory neuropathy type I. Am. J. Hum. Genet. 88, 99–105 10.1016/j.ajhg.2010.12.003 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
