

Vitamin C Sensitizes Melanoma to BET Inhibitors
- PMID: 29180474
- PMCID: PMC5771981
- DOI: 10.1158/0008-5472.CAN-17-2040
Vitamin C Sensitizes Melanoma to BET Inhibitors
Abstract
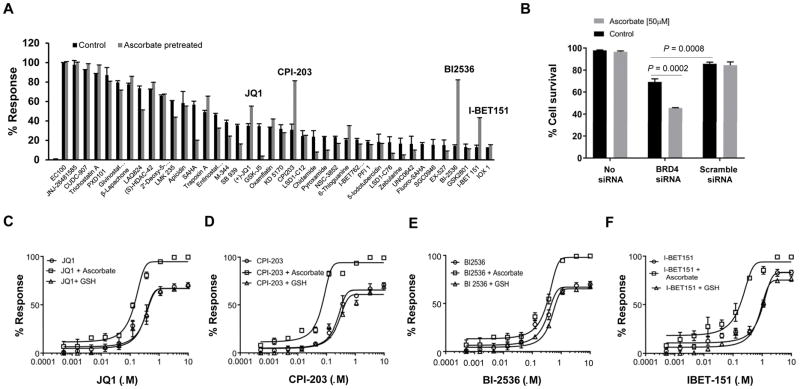
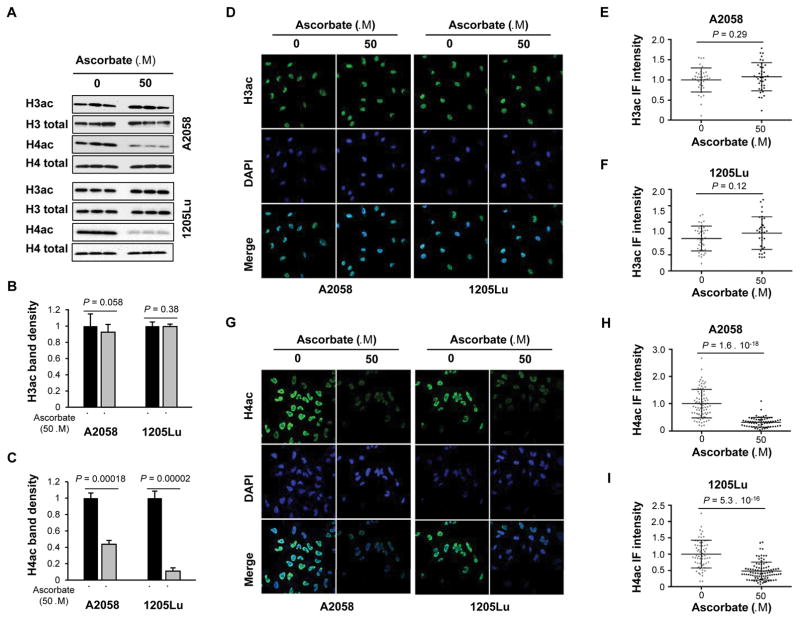
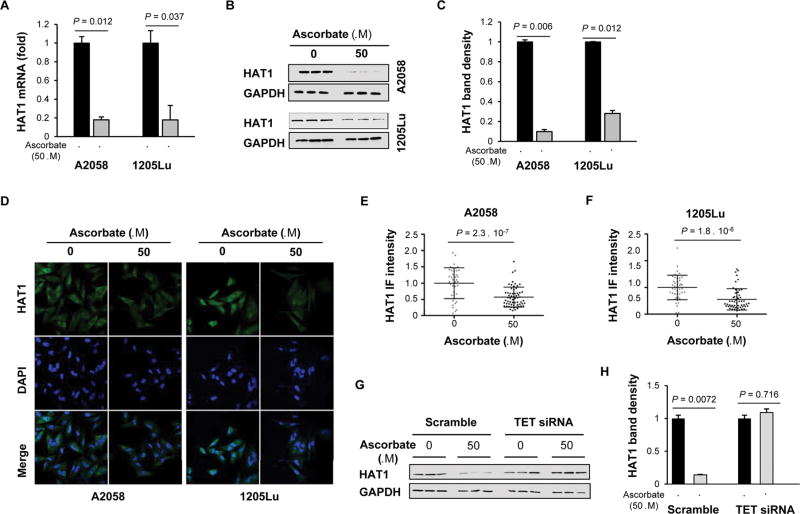
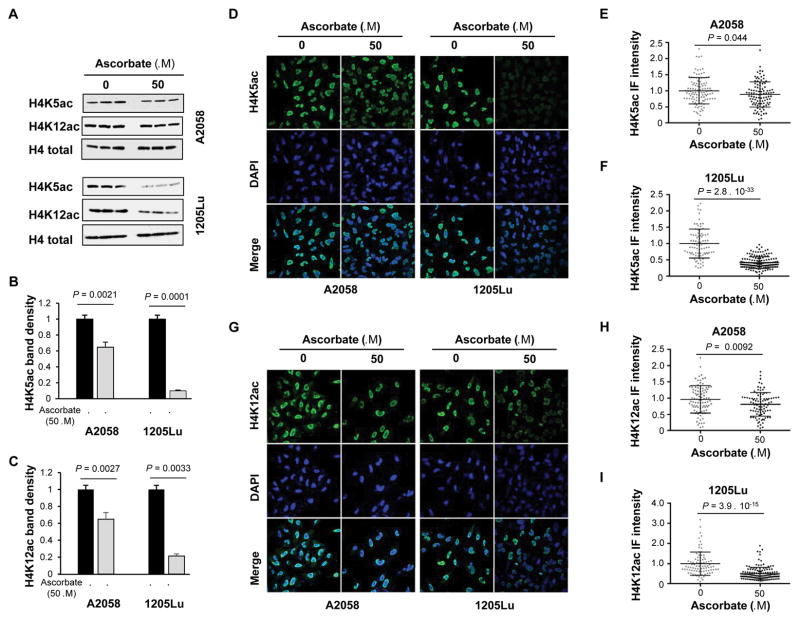
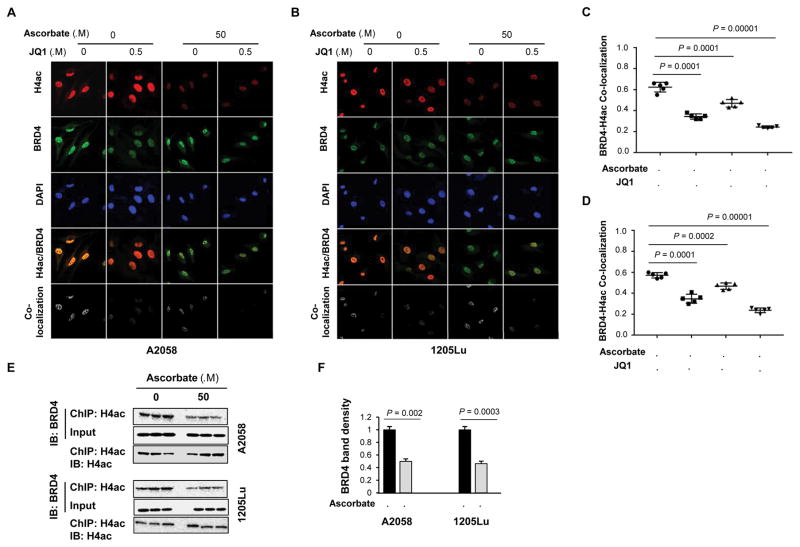
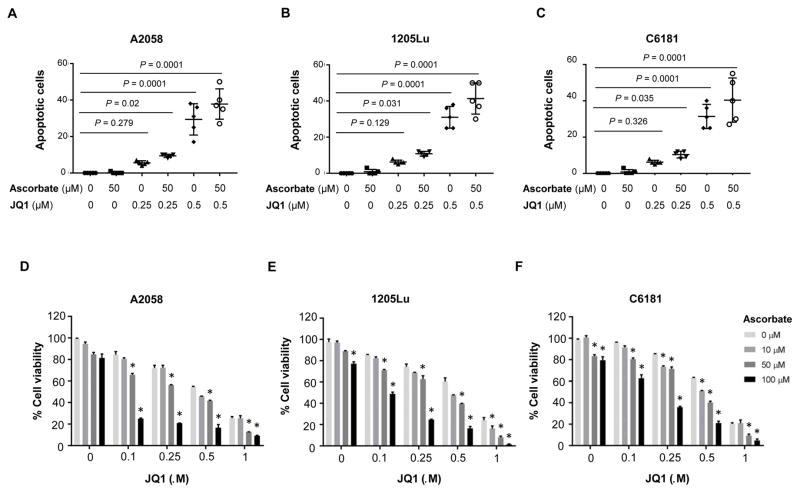
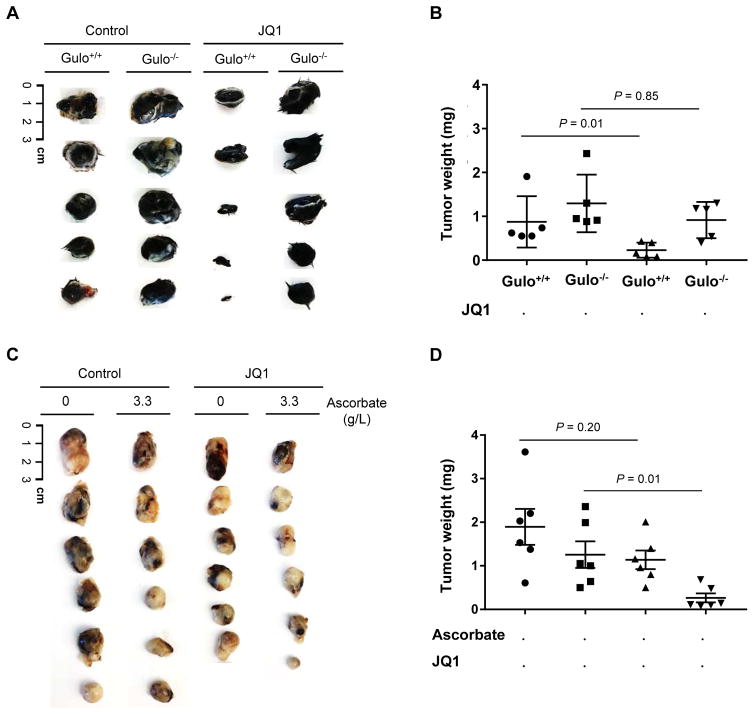
Bromodomain and extraterminal inhibitors (BETi) are promising cancer therapies, yet prominent side effects of BETi at effective doses have been reported in phase I clinical trials. Here, we screened a panel of small molecules targeting epigenetic modulators against human metastatic melanoma cells. Cells were pretreated with or without ascorbate (vitamin C), which promotes DNA demethylation and subsequently changes the sensitivity to drugs. Top hits were structurally unrelated BETi, including JQ1, I-BET151, CPI-203, and BI-2536. Ascorbate enhanced the efficacy of BETi by decreasing acetylation of histone H4, but not H3, while exerting no effect on the expression of BRD proteins. Histone acetyltransferase 1 (HAT1), which catalyzes H4K5ac and H4K12ac, was downregulated by ascorbate mainly via the TET-mediated DNA hydroxymethylation pathway. Loss of H4ac, especially H4K5ac and H4K12ac, disrupted the interaction between BRD4 and H4 by which ascorbate and BETi blocked the binding of BRD4 to acetylated histones. Cotreatment with ascorbate and JQ1 induced apoptosis and inhibited proliferation of cultured melanoma cells. Ascorbate deficiency as modeled in Gulo-/- mice diminished the treatment outcome of JQ1 for melanoma tumorgraft. In contrast, ascorbate supplementation lowered the effective dose of JQ1 needed to successfully inhibit melanoma tumors in mice. On the basis of our findings, future clinical trials with BETi should consider ascorbate levels in patients. Furthermore, ascorbate supplementation might help reduce the severe side effects that arise from BETi therapy by reducing the dosage necessary for treatment.Significance: This study shows that ascorbate can enhance the efficacy of BET inhibitors, providing a possible clinical solution to challenges arising in phase I trials from the dose-dependent side effects of this class of epigenetic therapy. Cancer Res; 78(2); 572-83. ©2017 AACR.
©2017 American Association for Cancer Research.
Conflict of interest statement
Figures
References
-
- Jain AK, Barton MC. Bromodomain Histone Readers and Cancer. J Mol Biol. 2017;429:2003–10. - PubMed
-
- Patino WD, Susa J. Epigenetics of cutaneous melanoma. Adv Dermatol. 2008;24:59–70. - PubMed
-
- Filippakopoulos P, Knapp S. Targeting bromodomains: epigenetic readers of lysine acetylation. Nat Rev Drug Discov. 2014;13:337–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
