

Proteomics of phosphorylation and protein dynamics during fertilization and meiotic exit in the Xenopus egg
- PMID: 29183978
- PMCID: PMC5740657
- DOI: 10.1073/pnas.1709207114
Proteomics of phosphorylation and protein dynamics during fertilization and meiotic exit in the Xenopus egg
Abstract
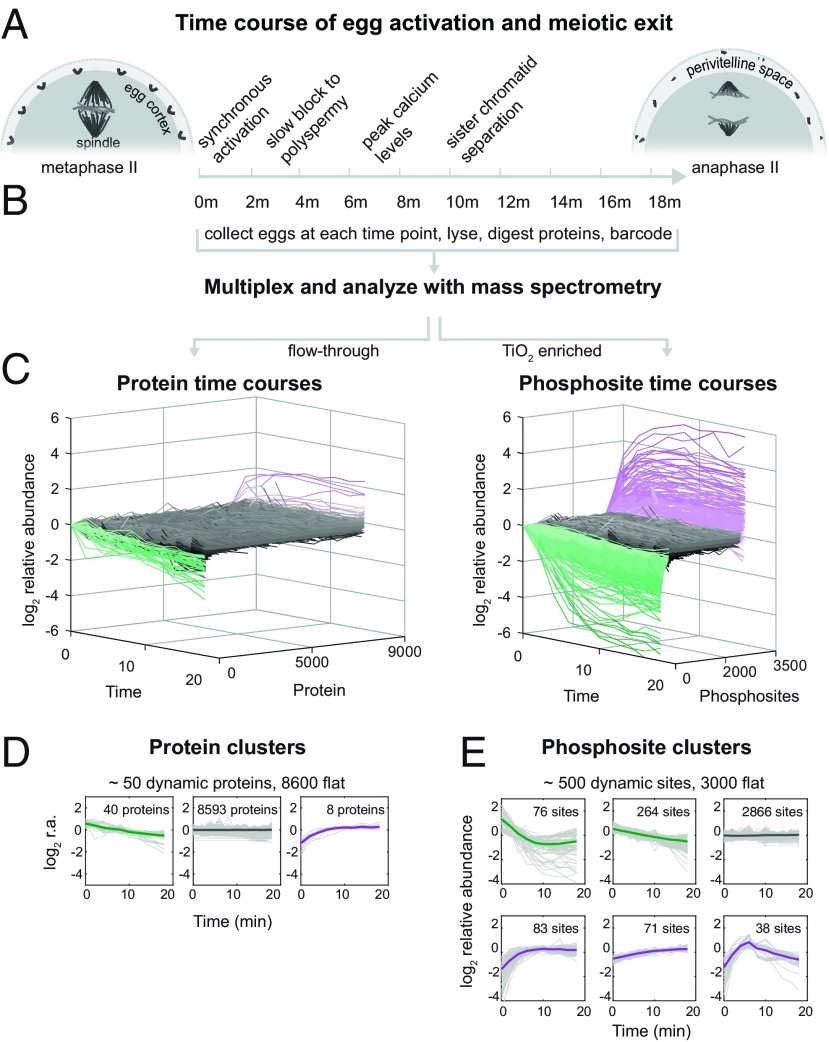
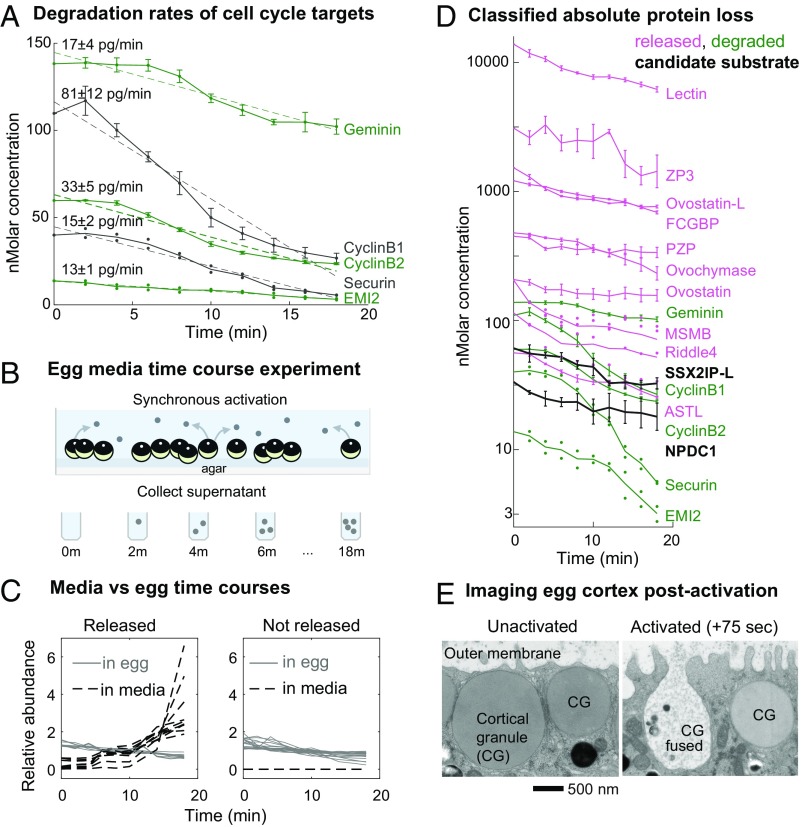
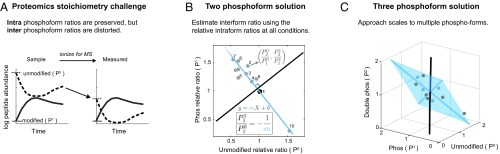
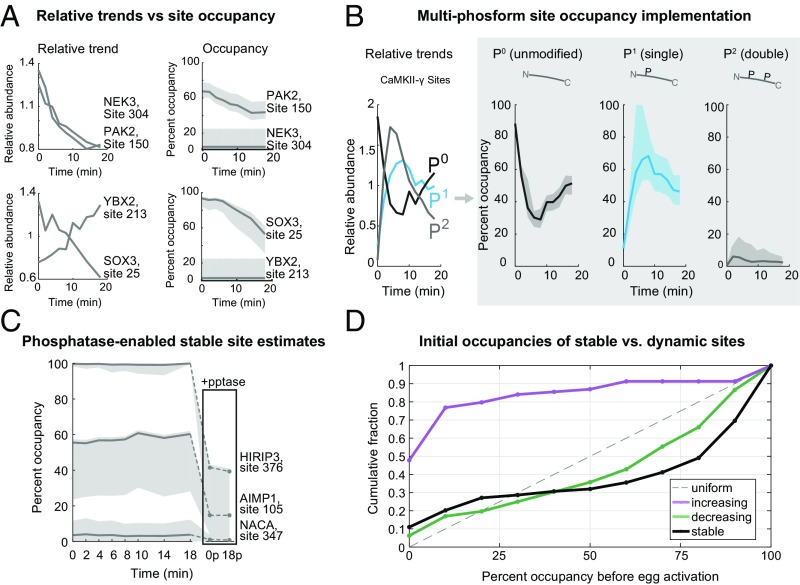
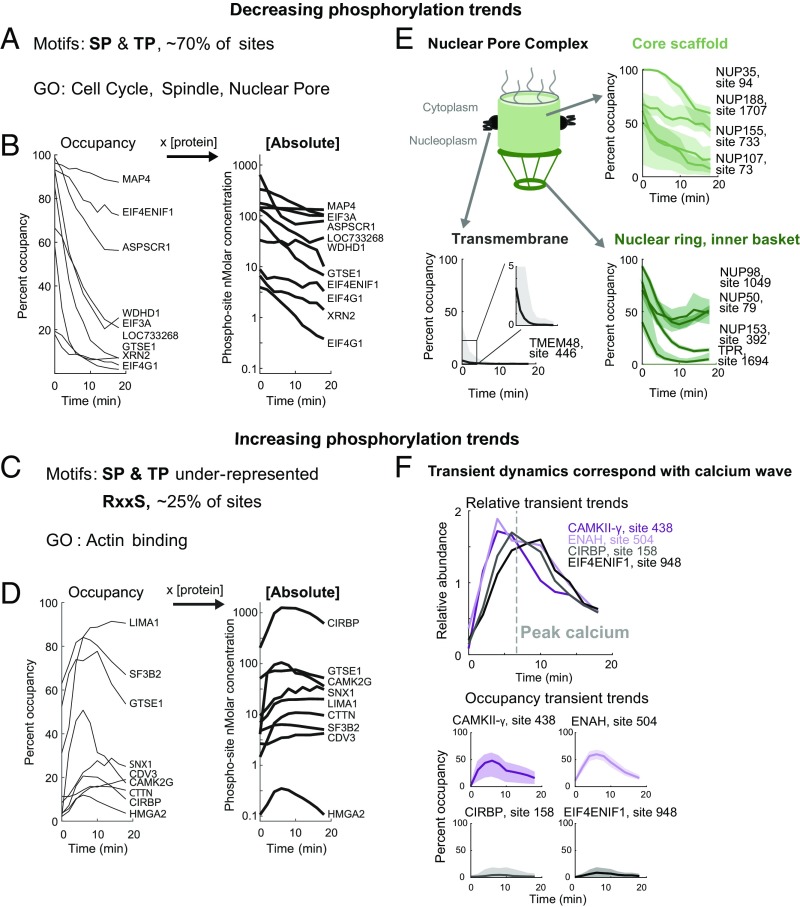
Fertilization releases the meiotic arrest and initiates the events that prepare the egg for the ensuing developmental program. Protein degradation and phosphorylation are known to regulate protein activity during this process. However, the full extent of protein loss and phosphoregulation is still unknown. We examined absolute protein and phosphosite dynamics of the fertilization response by mass spectrometry-based proteomics in electroactivated eggs. To do this, we developed an approach for calculating the stoichiometry of phosphosites from multiplexed proteomics that is compatible with dynamic, stable, and multisite phosphorylation. Overall, the data suggest that degradation is limited to a few low-abundance proteins. However, this degradation promotes extensive dephosphorylation that occurs over a wide range of abundances during meiotic exit. We also show that eggs release a large amount of protein into the medium just after fertilization, most likely related to the blocks to polyspermy. Concomitantly, there is a substantial increase in phosphorylation likely tied to calcium-activated kinases. We identify putative degradation targets and components of the slow block to polyspermy. The analytical approaches demonstrated here are broadly applicable to studies of dynamic biological systems.
Keywords: TMT; cell cycle; confidence intervals; polyspermy block; stoichiometry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
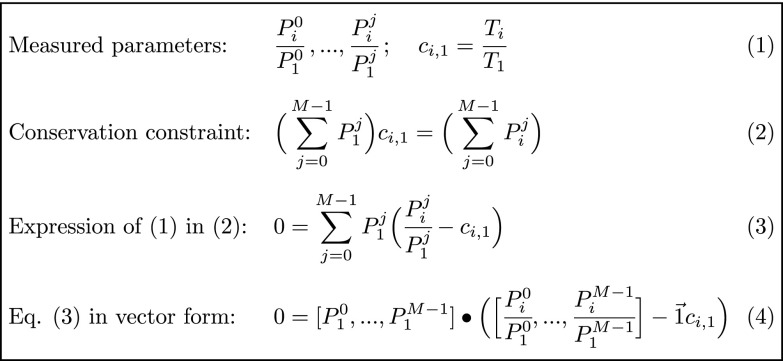
References
-
- Tunquist BJ, Maller JL. Under arrest: Cytostatic factor (CSF)-mediated metaphase arrest in vertebrate eggs. Genes Dev. 2003;17:683–710. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
