

Vasopressin excites interneurons to suppress hippocampal network activity across a broad span of brain maturity at birth
- PMID: 29183979
- PMCID: PMC5740624
- DOI: 10.1073/pnas.1717337114
Vasopressin excites interneurons to suppress hippocampal network activity across a broad span of brain maturity at birth
Abstract
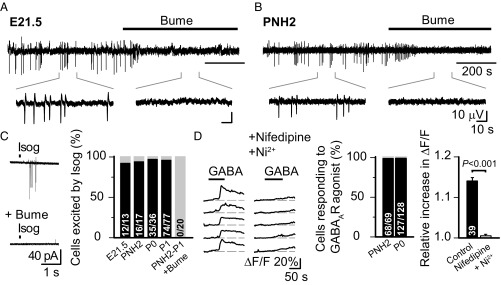
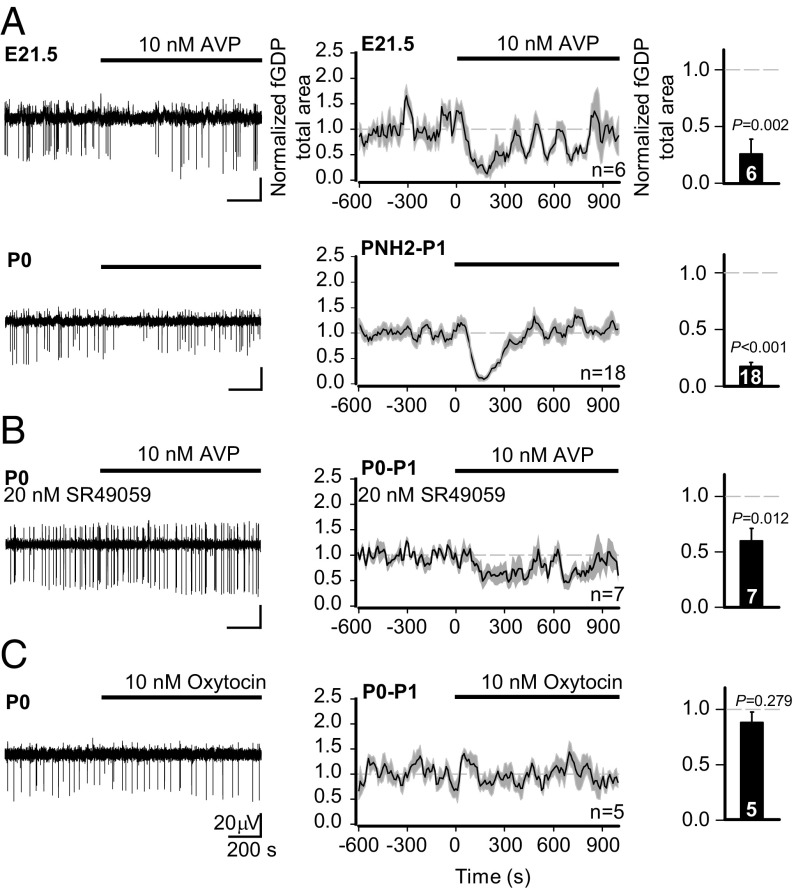
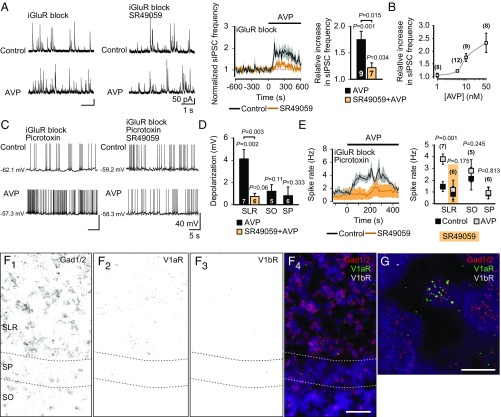
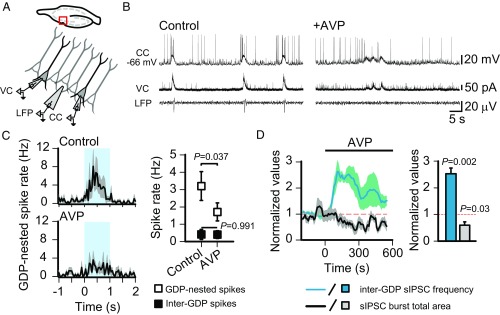
During birth in mammals, a pronounced surge of fetal peripheral stress hormones takes place to promote survival in the transition to the extrauterine environment. However, it is not known whether the hormonal signaling involves central pathways with direct protective effects on the perinatal brain. Here, we show that arginine vasopressin specifically activates interneurons to suppress spontaneous network events in the perinatal hippocampus. Experiments done on the altricial rat and precocial guinea pig neonate demonstrated that the effect of vasopressin is not dependent on the level of maturation (depolarizing vs. hyperpolarizing) of postsynaptic GABAA receptor actions. Thus, the fetal mammalian brain is equipped with an evolutionarily conserved mechanism well-suited to suppress energetically expensive correlated network events under conditions of reduced oxygen supply at birth.
Keywords: GDP; KCC2; birth asphyxia; bumetanide; oxytocin.
Copyright © 2017 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Liu L, et al. Global, regional, and national causes of child mortality in 2000-13, with projections to inform post-2015 priorities: An updated systematic analysis. Lancet. 2015;385:430–440. - PubMed
-
- Motoyama EK, et al. Effect of cortisol on the maturation of fetal rabbit lungs. Pediatrics. 1971;48:547–555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
