

Deubiquitylase USP9X suppresses tumorigenesis by stabilizing large tumor suppressor kinase 2 (LATS2) in the Hippo pathway
- PMID: 29183995
- PMCID: PMC5787797
- DOI: 10.1074/jbc.RA117.000392
Deubiquitylase USP9X suppresses tumorigenesis by stabilizing large tumor suppressor kinase 2 (LATS2) in the Hippo pathway
Abstract
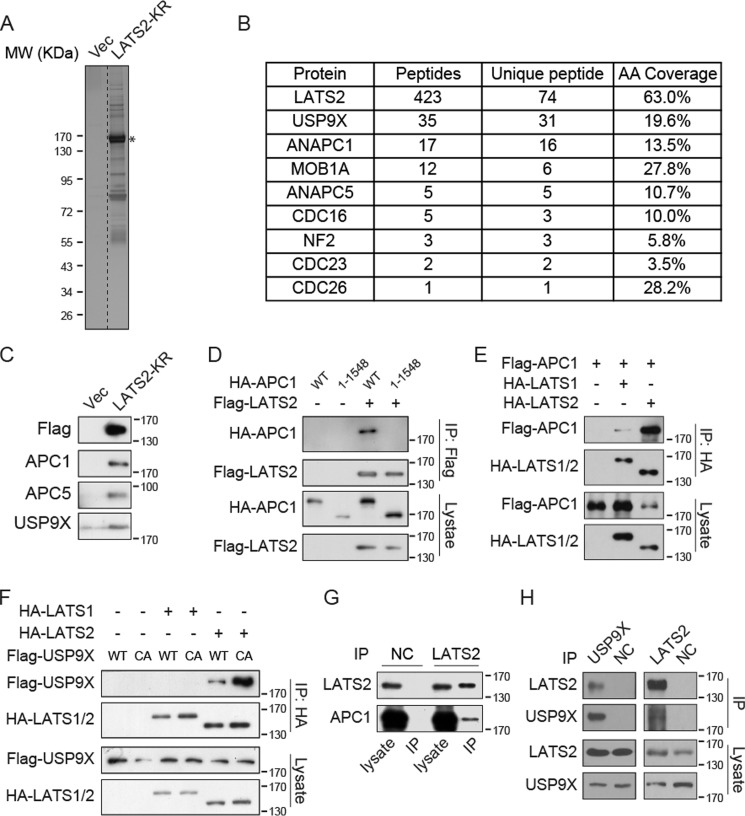
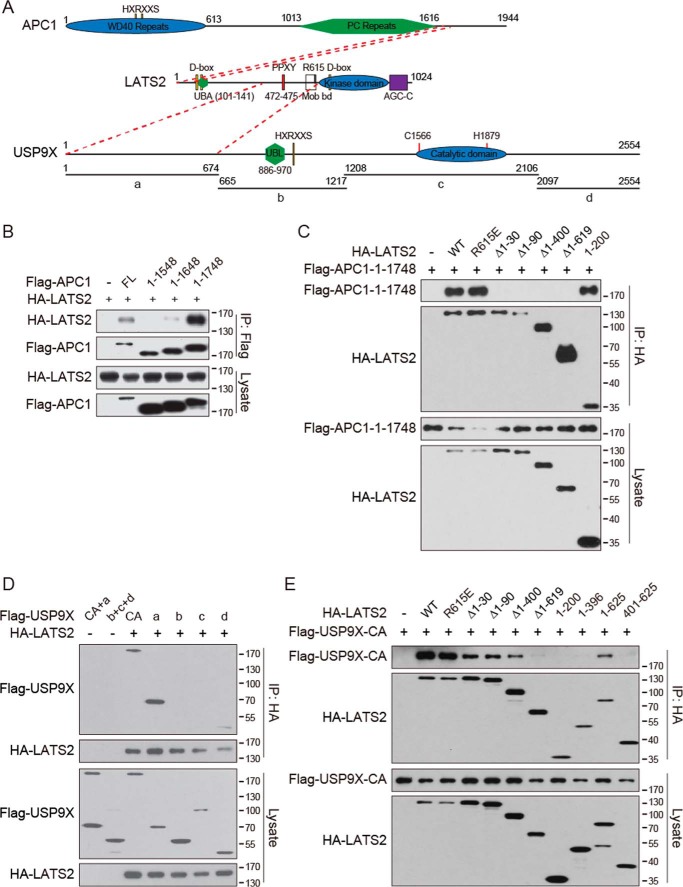
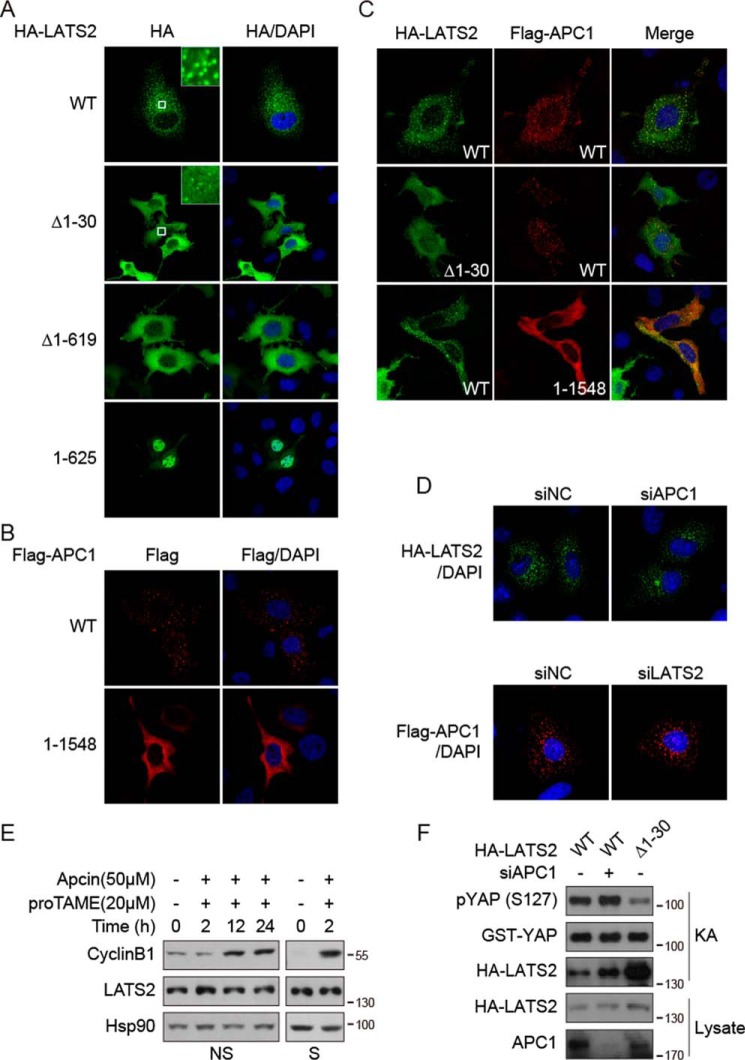
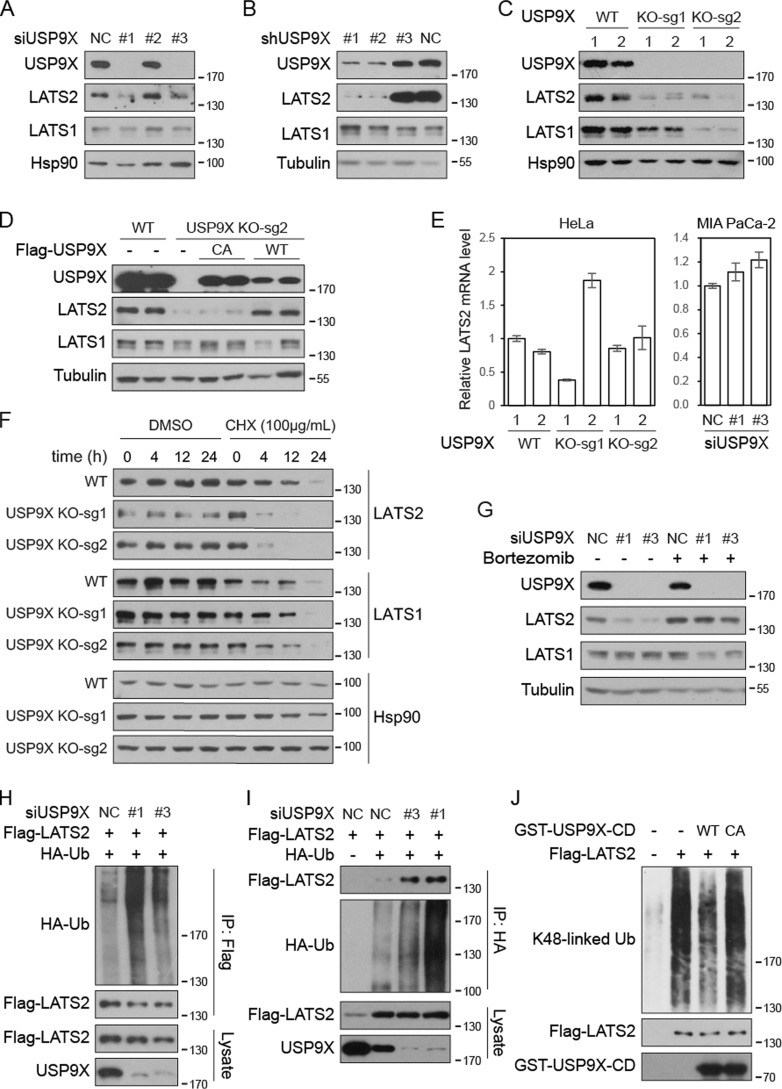
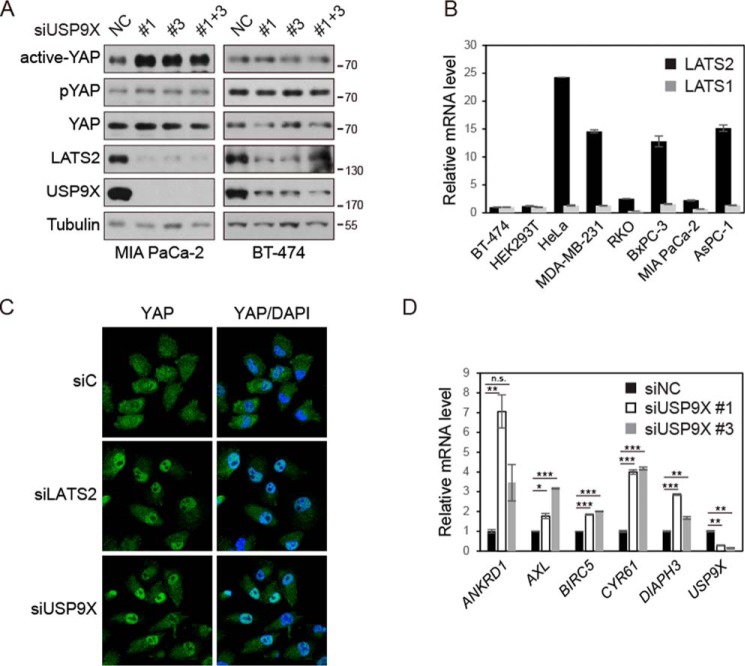
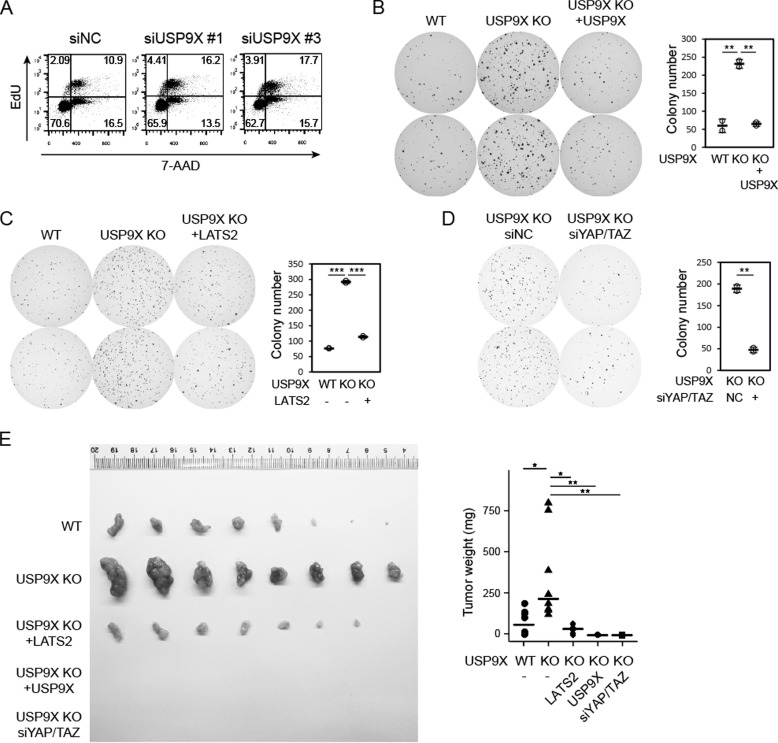
The Hippo pathway plays important roles in controlling organ size and in suppressing tumorigenesis through large tumor suppressor kinase 1/2 (LATS1/2)-mediated phosphorylation of YAP/TAZ transcription co-activators. The kinase activity of LATS1/2 is regulated by phosphorylation in response to extracellular signals. Moreover, LATS2 protein levels are repressed by the ubiquitin-proteasome system in conditions such as hypoxia. However, the mechanism that removes the ubiquitin modification from LATS2 and thereby stabilizes the protein is not well understood. Here, using tandem affinity purification (TAP), we found that anaphase-promoting complex/cyclosome (APC/C), a ubiquitin ligase complex, and USP9X, a deubiquitylase, specifically interact with LATS2. We also found that although APC1 co-localizes with LATS2 to intracellular vesicle structures, it does not regulate LATS2 protein levels and activity. In contrast, USP9X ablation drastically diminished LATS2 protein levels. We further demonstrated that USP9X deubiquitinates LATS2 and thus prevents LATS2 degradation by the proteasome. Furthermore, in pancreatic cancer cells, USP9X loss activated YAP and enhanced the oncogenic potential of the cells. In addition, the tumorigenesis induced by the USP9X ablation depended not only on LATS2 repression, but also on YAP/TAZ activity. We conclude that USP9X is a deubiquitylase of the Hippo pathway kinase LATS2 and that the Hippo pathway functions as a downstream signaling cascade that mediates USP9X's tumor-suppressive activity.
Keywords: Hippo pathway; LATS2; USP9X; anaphase-promoting complex; deubiquitination; protein stability; signal transduction; tumor suppressor gene; ubiquitination.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Deubiquitinating Enzyme USP9X Suppresses Tumor Growth via LATS Kinase and Core Components of the Hippo Pathway.Cancer Res. 2017 Sep 15;77(18):4921-4933. doi: 10.1158/0008-5472.CAN-16-3413. Epub 2017 Jul 18. Cancer Res. 2017. PMID: 28720576 Free PMC article.
-
Hypoxia regulates Hippo signalling through the SIAH2 ubiquitin E3 ligase.Nat Cell Biol. 2015 Jan;17(1):95-103. doi: 10.1038/ncb3073. Epub 2014 Dec 1. Nat Cell Biol. 2015. PMID: 25438054
-
Tumor suppressor LATS1 is a negative regulator of oncogene YAP.J Biol Chem. 2008 Feb 29;283(9):5496-509. doi: 10.1074/jbc.M709037200. Epub 2007 Dec 24. J Biol Chem. 2008. PMID: 18158288
-
The LATS1 and LATS2 tumor suppressors: beyond the Hippo pathway.Cell Death Differ. 2017 Sep;24(9):1488-1501. doi: 10.1038/cdd.2017.99. Epub 2017 Jun 23. Cell Death Differ. 2017. PMID: 28644436 Free PMC article. Review.
-
Hippo-YAP/TAZ signaling in angiogenesis.BMB Rep. 2018 Mar;51(3):157-162. doi: 10.5483/bmbrep.2018.51.3.016. BMB Rep. 2018. PMID: 29366443 Free PMC article. Review.
Cited by
-
Treating ARID1A mutated cancers by harnessing synthetic lethality and DNA damage response.J Biomed Sci. 2022 Sep 19;29(1):71. doi: 10.1186/s12929-022-00856-5. J Biomed Sci. 2022. PMID: 36123603 Free PMC article. Review.
-
Overexpression of the Ubiquitin-Specific Peptidase 9 X-Linked (USP9X) Gene is Associated with Upregulation of Cyclin D1 (CCND1) and Downregulation of Cyclin-Dependent Inhibitor Kinase 1A (CDKN1A) in Breast Cancer Tissue and Cell Lines.Med Sci Monit. 2019 Jun 6;25:4207-4216. doi: 10.12659/MSM.914742. Med Sci Monit. 2019. PMID: 31169265 Free PMC article.
-
The E3 ubiquitin ligase, FBXW5, promotes the migration and invasion of gastric cancer through the dysregulation of the Hippo pathway.Cell Death Discov. 2022 Feb 24;8(1):79. doi: 10.1038/s41420-022-00868-y. Cell Death Discov. 2022. PMID: 35210431 Free PMC article.
-
Precise Identification of Recurrent Somatic Mutations in Oral Cancer Through Whole-Exome Sequencing Using Multiple Mutation Calling Pipelines.Front Oncol. 2021 Nov 29;11:741626. doi: 10.3389/fonc.2021.741626. eCollection 2021. Front Oncol. 2021. PMID: 34912705 Free PMC article.
-
Myricetin Suppresses the Propagation of Hepatocellular Carcinoma via Down-Regulating Expression of YAP.Cells. 2019 Apr 17;8(4):358. doi: 10.3390/cells8040358. Cells. 2019. PMID: 30999669 Free PMC article.
References
-
- Song H., Mak K. K., Topol L., Yun K., Hu J., Garrett L., Chen Y., Park O., Chang J., Simpson R. M., Wang C. Y., Gao B., Jiang J., and Yang Y. (2010) Mammalian Mst1 and Mst2 kinases play essential roles in organ size control and tumor suppression. Proc. Natl. Acad. Sci. U.S.A. 107, 1431–1436 10.1073/pnas.0911409107 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
