

Zinc Limitation Induces a Hyper-Adherent Goliath Phenotype in Candida albicans
- PMID: 29184547
- PMCID: PMC5694484
- DOI: 10.3389/fmicb.2017.02238
Zinc Limitation Induces a Hyper-Adherent Goliath Phenotype in Candida albicans
Abstract
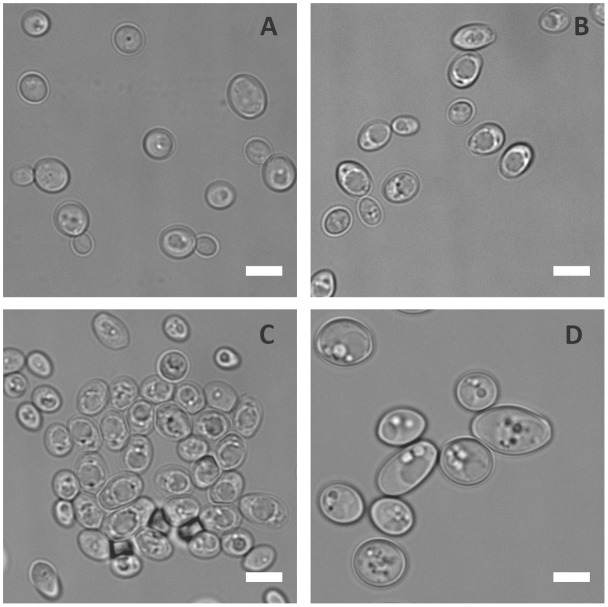
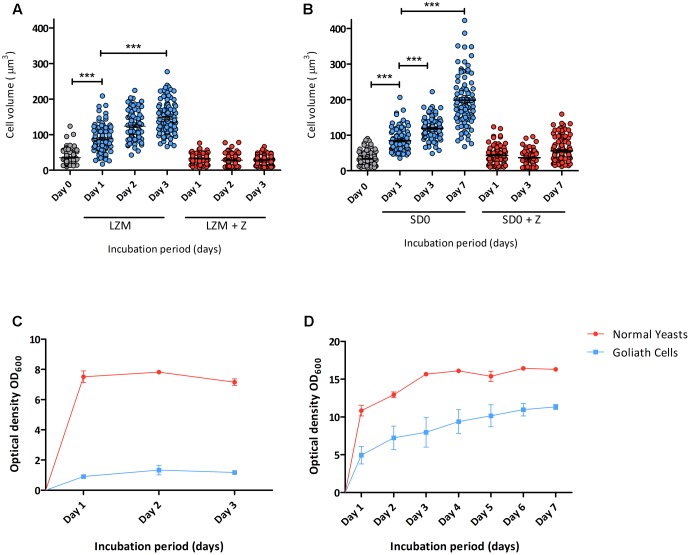
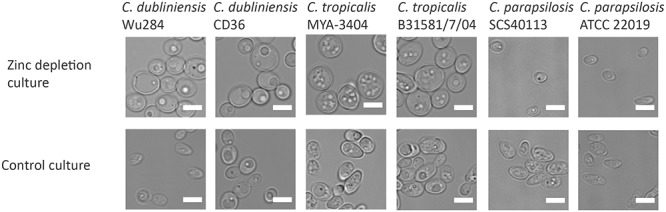
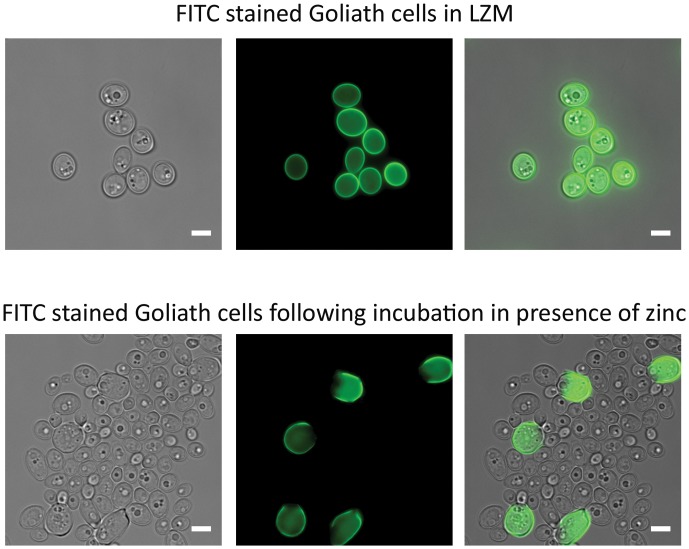
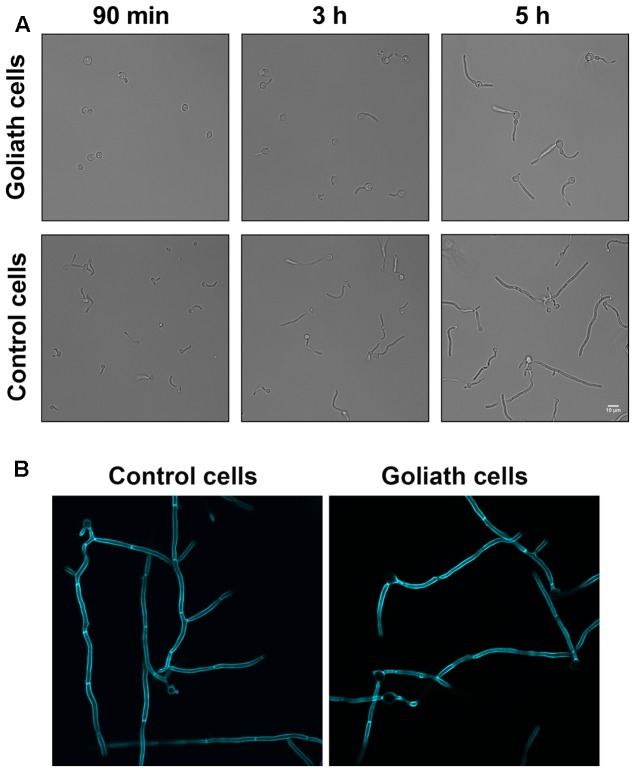
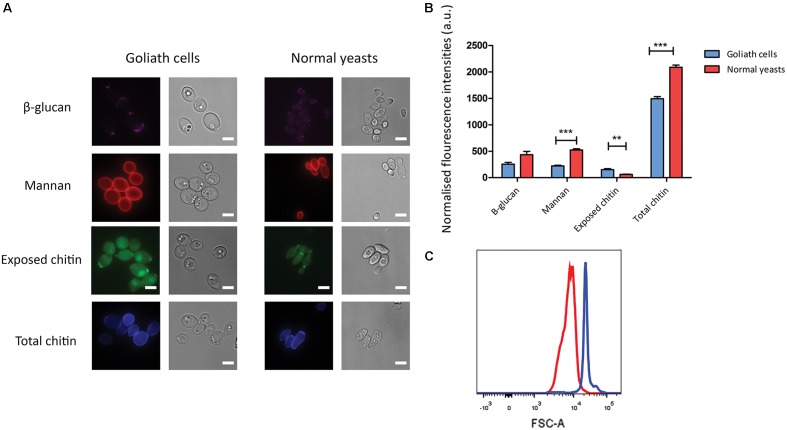
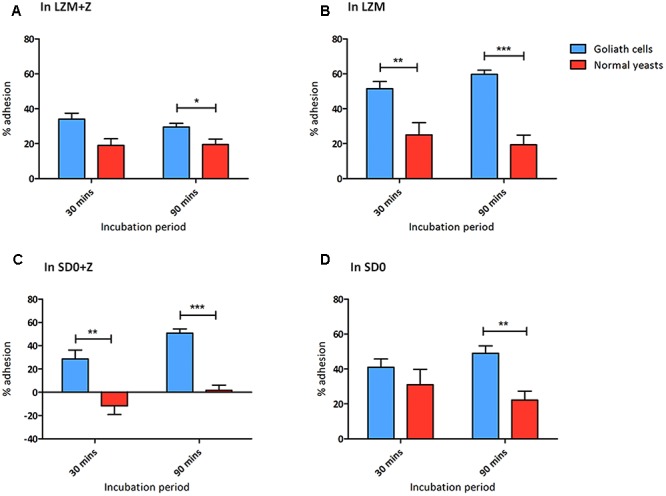
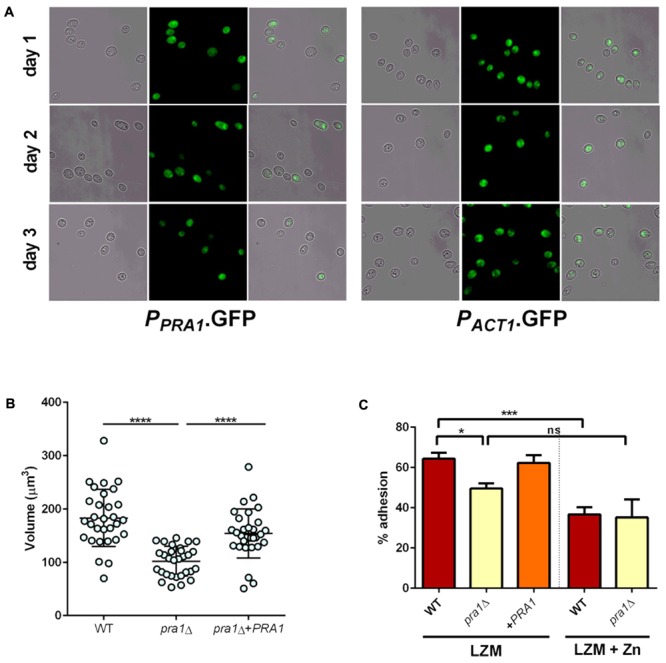
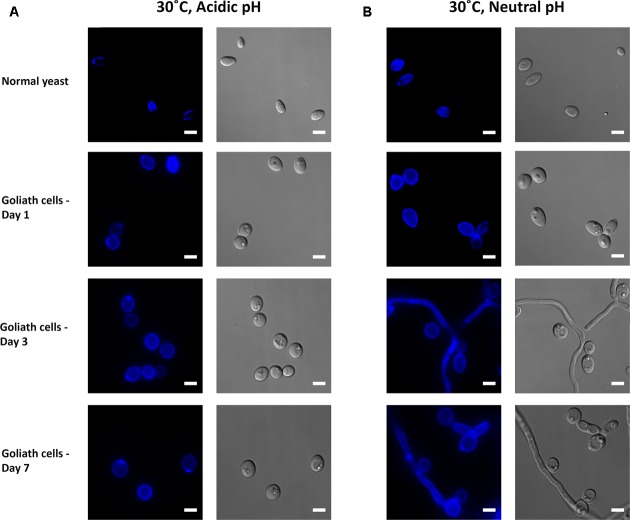
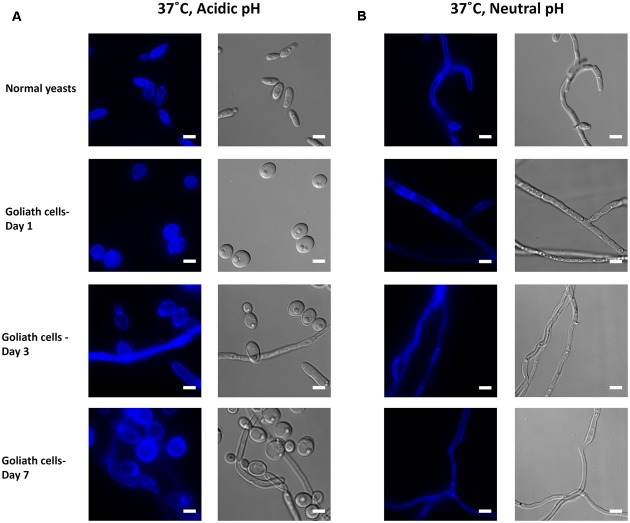
Pathogenic microorganisms often face acute micronutrient limitation during infection due to the action of host-mediated nutritional immunity. The human fungal pathogen Candida albicans is polymorphic and its morphological plasticity is one of its most widely recognized pathogenicity attributes. Here we investigated the effect of zinc, iron, manganese, and copper limitation on C. albicans morphology. Restriction of zinc specifically resulted in the formation of enlarged, spherical yeasts, a phenotype which we term Goliath cells. This cellular response to zinc restriction was conserved in C. albicans, C. dubliniensis and C. tropicalis, but not in C. parapsilosis, C. lusitaniae or Debaryomyces hansenii, suggesting that it may have emerged in the last common ancestor of these related pathogenic species. Cell wall analysis revealed proportionally more chitin exposure on the Goliath cell surface. Importantly, these cells were hyper-adherent, suggesting a possible role in pathogenicity. Interestingly, the zincophore-encoding gene PRA1 was expressed by Goliath cells in zinc limited media and lack of Pra1 inhibited both cellular enlargement and adhesion. Goliath cells represent a further layer of Candida phenotypic plasticity.
Keywords: Candida albicans; adhesion; fungal pathogen; micronutrients; morphogenesis; morphology; nutritional immunity; zinc.
Figures

References
-
- Alasio T. M., Lento P. A., Bottone E. J. (2003). Giant blastoconidia of Candida albicans. A case report and review of the literature. Arch. Pathol. Lab. Med. 127 868–871. - PubMed
-
- Ameglio F., Di Giorgio C., Terzaroli P., Gandolfo G. M. (1990). Giant cell production by C. albicans cultured in xylitol. Microbiologica 13 343–346. - PubMed
-
- Amich J., Vicentefranqueira R., Mellado E., Ruiz-Carmuega A., Leal F., Calera J. A. (2014). The ZrfC alkaline zinc transporter is required for Aspergillus fumigatus virulence and its growth in the presence of the Zn/Mn-chelating protein calprotectin. Cell Microbiol. 16 548–564. 10.1111/cmi.12238 - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources