

Internalization of the Active Subunit of the Aggregatibacter actinomycetemcomitans Cytolethal Distending Toxin Is Dependent upon Cellugyrin (Synaptogyrin 2), a Host Cell Non-Neuronal Paralog of the Synaptic Vesicle Protein, Synaptogyrin 1
- PMID: 29184850
- PMCID: PMC5694546
- DOI: 10.3389/fcimb.2017.00469
Internalization of the Active Subunit of the Aggregatibacter actinomycetemcomitans Cytolethal Distending Toxin Is Dependent upon Cellugyrin (Synaptogyrin 2), a Host Cell Non-Neuronal Paralog of the Synaptic Vesicle Protein, Synaptogyrin 1
Abstract
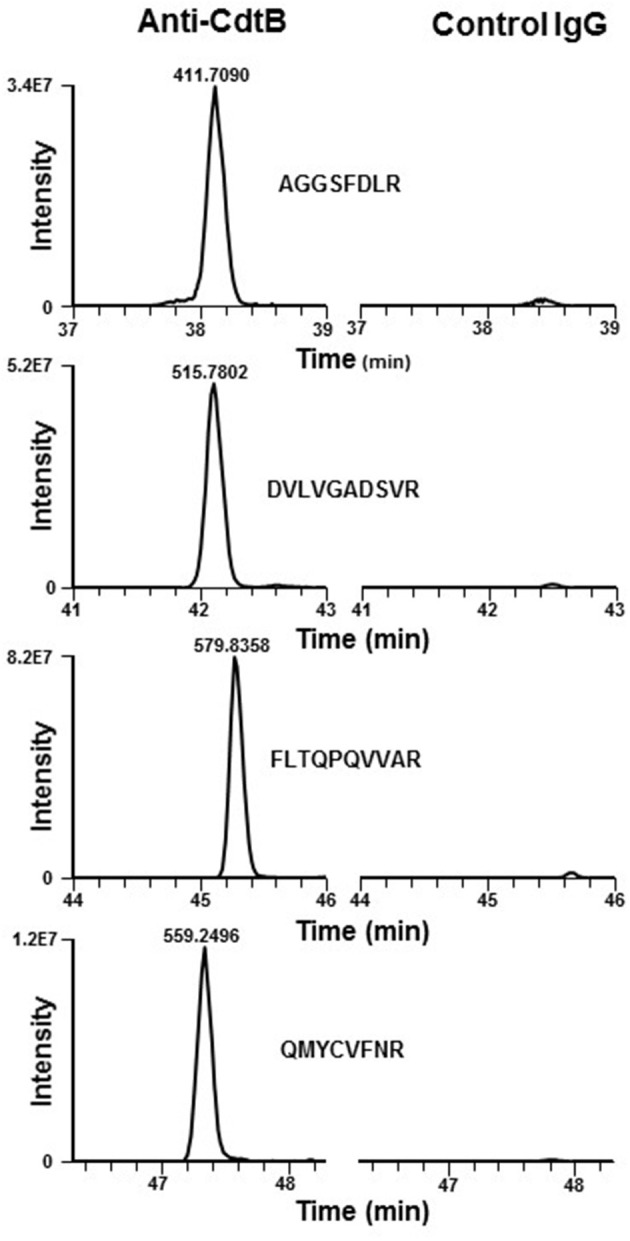
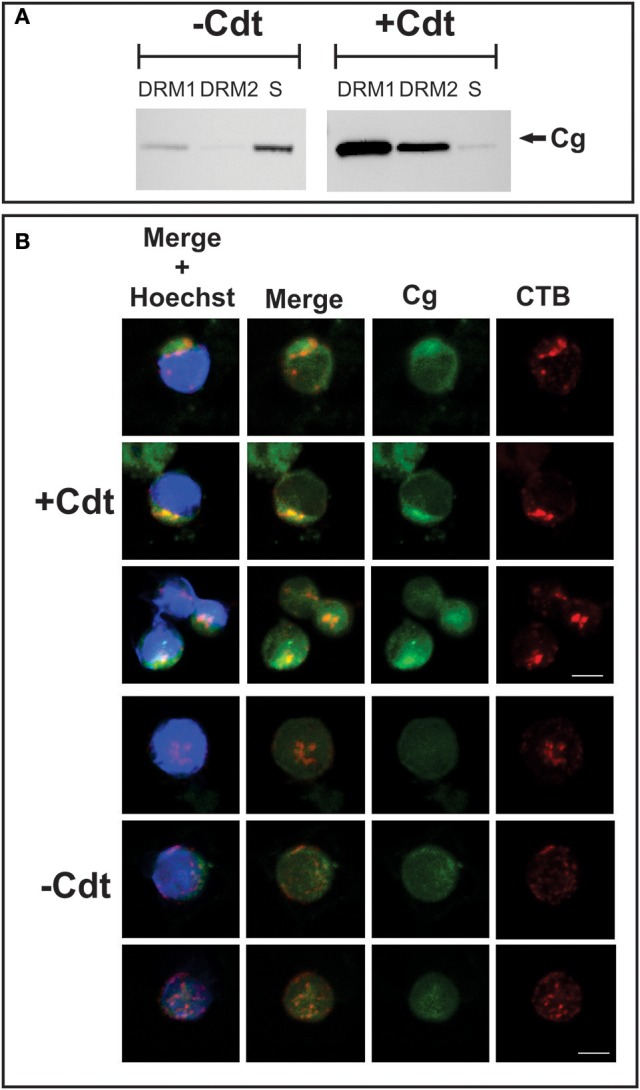
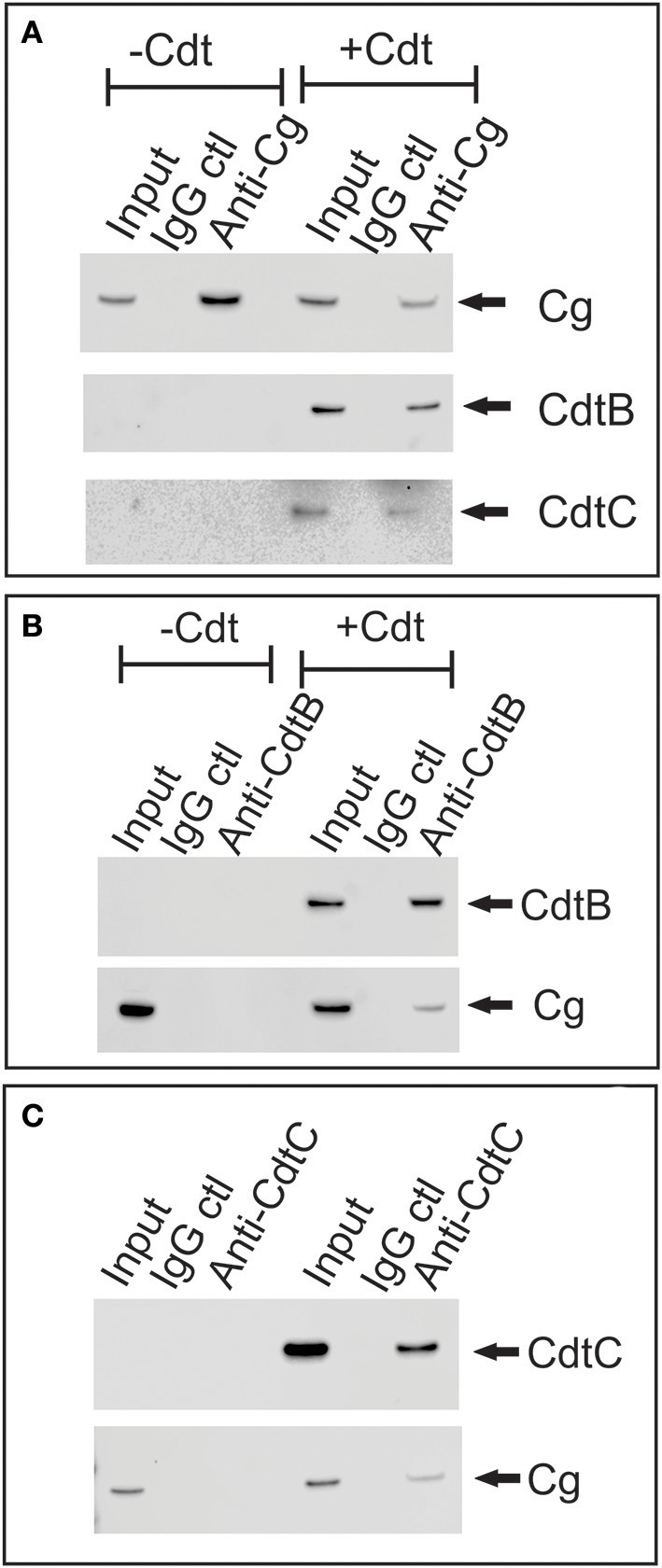
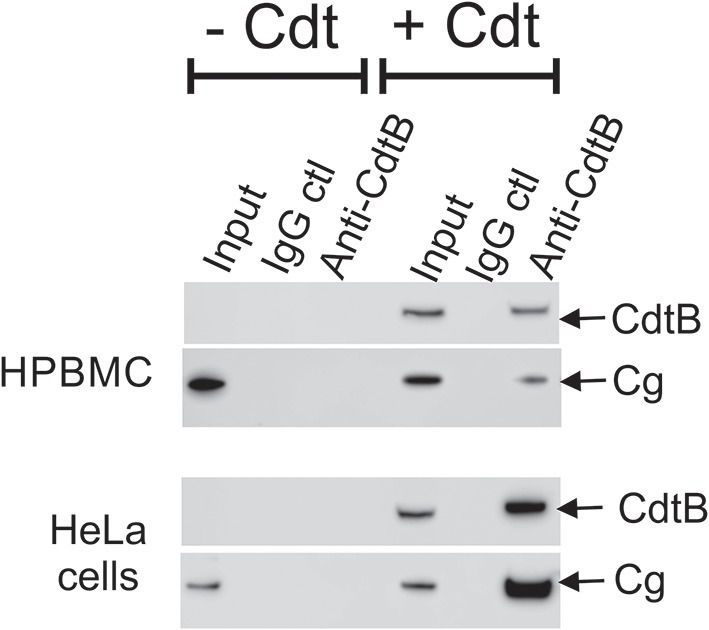
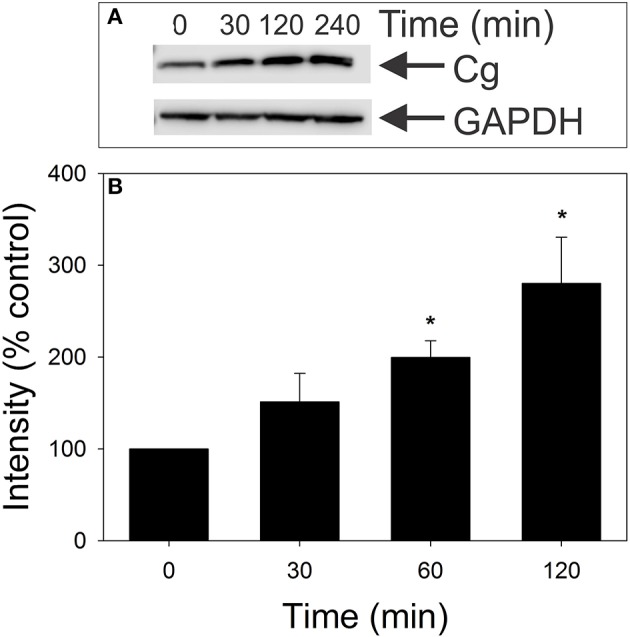
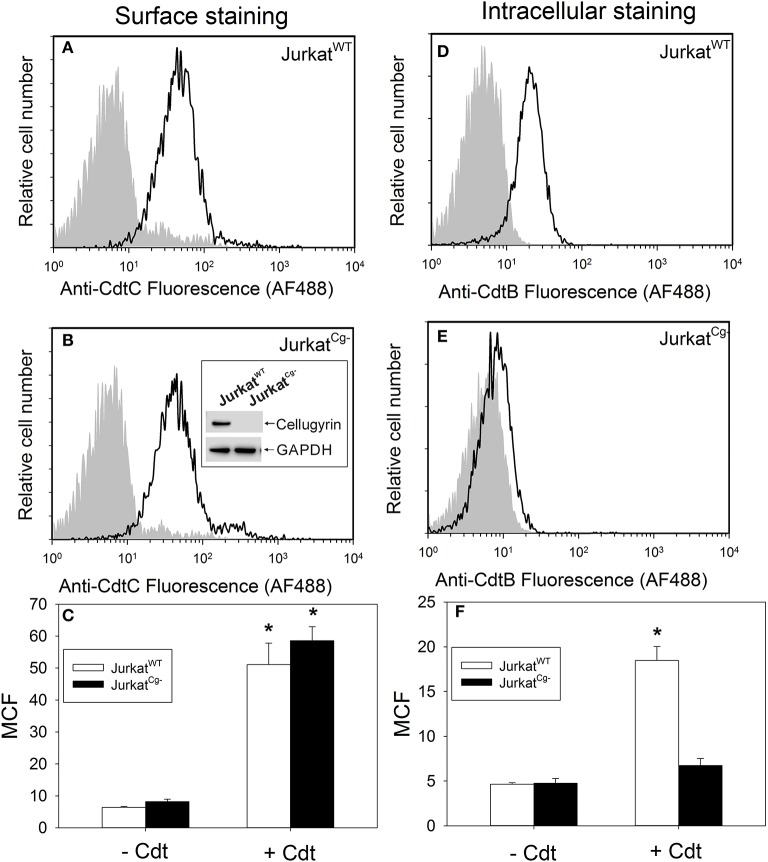
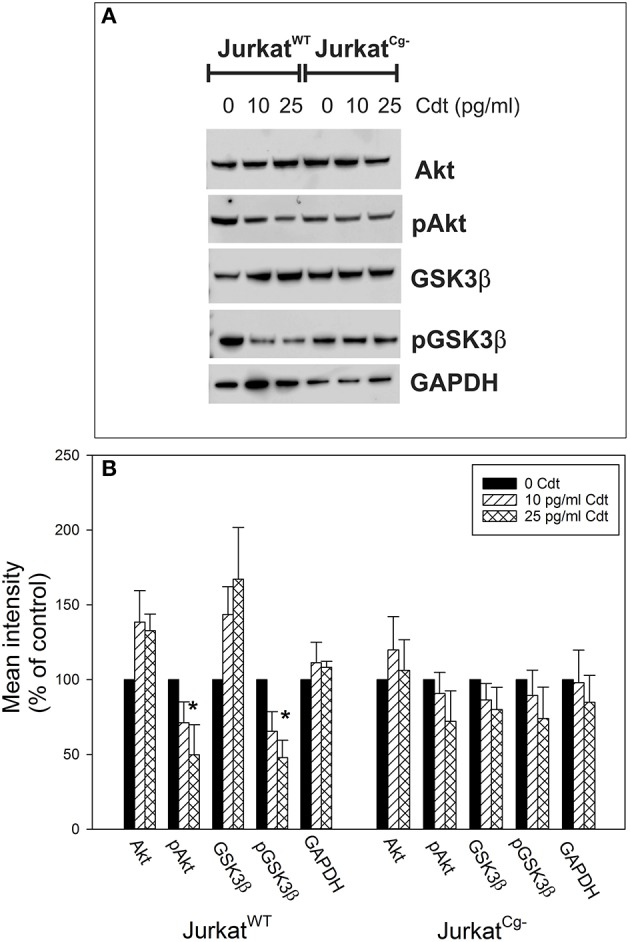
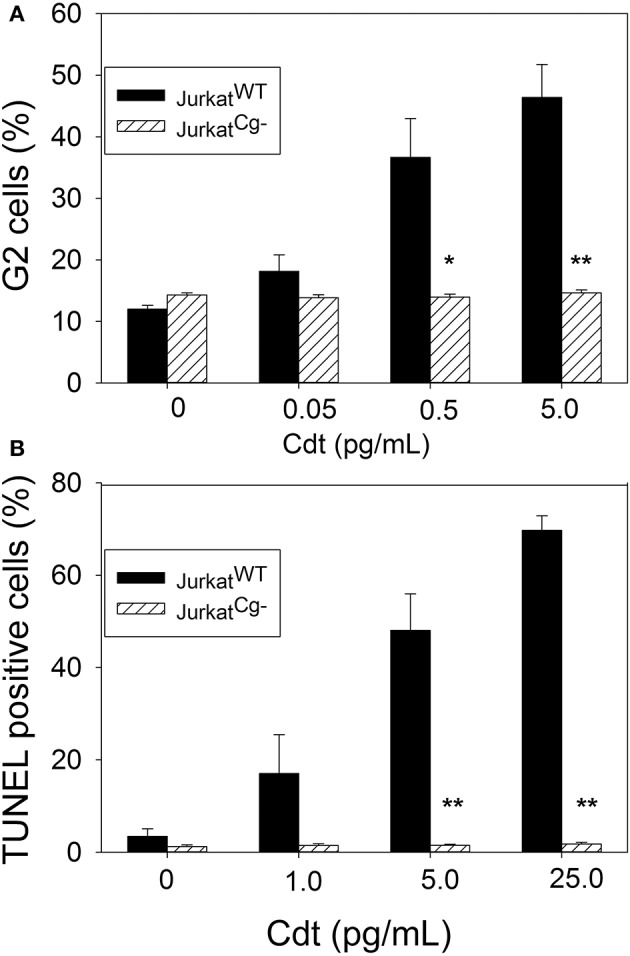
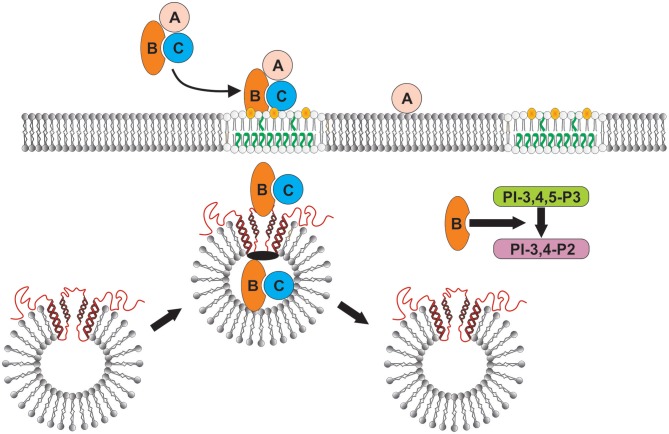
The Aggregatibacter actinomycetemcomitans cytolethal distending toxin (Cdt) is a heterotrimeric AB2 toxin capable of inducing lymphocytes, and other cell types, to undergo cell cycle arrest and apoptosis. Exposure to Cdt results in binding to the cell surface followed by internalization and translocation of the active subunit, CdtB, to intracellular compartments. These events are dependent upon toxin binding to cholesterol in the context of lipid rich membrane microdomains often referred to as lipid rafts. We now demonstrate that, in addition to binding to the plasma membrane of lymphocytes, another early and critical event initiated by Cdt is the translocation of the host cell protein, cellugyrin (synaptogyrin-2) to the same cholesterol-rich microdomains. Furthermore, we demonstrate that cellugyrin is an intracellular binding partner for CdtB as demonstrated by immunoprecipitation. Using CRISPR/cas9 gene editing we established a Jurkat cell line deficient in cellugyrin expression (JurkatCg-); these cells were capable of binding Cdt, but unable to internalize CdtB. Furthermore, JurkatCg- cells were not susceptible to Cdt-induced toxicity; these cells failed to exhibit blockade of the PI-3K signaling pathway, cell cycle arrest or cell death. We propose that cellugyrin plays a critical role in the internalization and translocation of CdtB to critical intracellular target sites. These studies provide critical new insight into the mechanism by which Cdt, and in particular, CdtB is able to induce toxicity.
Keywords: Aggregatibacter actinomycetemcomitans; apoptosis; cell cycle arrest; cytolethal distending toxin; lymphocytes; pathogenesis; toxin.
Figures
References
-
- Bielaszewska M., Ruter C., Bauwens A., Greune L., Jarosch K. A., Steil D., et al. (2017). Host cell interactions of outer membrane vesicle-associated virulence factors of enterohemorrhagic Escherichia coli O157: intracellular delivery, trafficking and mechanisms of cell injury. PLoS Pathog. 13:e1006159. 10.1371/journal.ppat.1006159 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous