

Construction of PRDM9 allele-specific recombination maps in cattle using large-scale pedigree analysis and genome-wide single sperm genomics
- PMID: 29186399
- PMCID: PMC5909443
- DOI: 10.1093/dnares/dsx048
Construction of PRDM9 allele-specific recombination maps in cattle using large-scale pedigree analysis and genome-wide single sperm genomics
Abstract
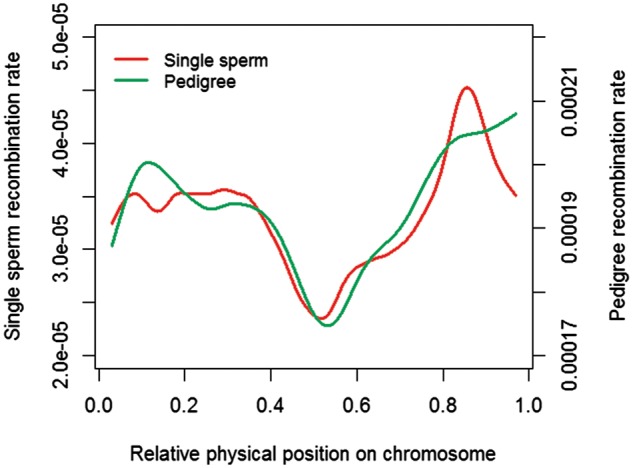
PRDM9 contributes to hybrid sterility and species evolution. However, its role is to be confirmed in cattle, a major domesticated livestock species. We previously found an association near PRDM9 with cattle recombination features, but the causative variants are still unknown. Using millions of genotyped cattle with pedigree information, we characterized five PRDM9 alleles and generated allele-specific recombination maps. By examining allele-specific recombination patterns, we observed the impact of PRDM9 on global distribution of recombination, especially in the two ends of chromosomes. We also showed strong associations between recombination hotspot regions and functional mutations within PRDM9 zinc finger domain. More importantly, we found one allele of PRDM9 to be very different from others in both protein composition and recombination landscape, indicating the causative role of this allele on the association between PRDM9 and cattle recombination. When comparing recombination maps from sperm and pedigree data, we observed similar genome-wide recombination patterns, validating the quality of pedigree-based results. Collectively, these evidence supported PRDM9 alleles as causal variants for the reported association with cattle recombination. Our study comprehensively surveyed the bovine PRDM9 alleles, generated allele-specific recombination maps, and expanded our understanding of the role of PRDM9 on genome distribution of recombination.
Figures



References
-
- Barton N.H., Charlesworth B.. 1998, Why sex and recombination?, Science, 281, 1986–90. - PubMed
-
- Stumpf M.P., McVean G.A.. 2003, Estimating recombination rates from population-genetic data, Nat. Rev. Genet., 4, 959–68. - PubMed
-
- Kauppi L., Jeffreys A.J., Keeney S.. 2004, Where the crossovers are: recombination distributions in mammals, Nat. Rev. Genet., 5, 413–24. - PubMed
-
- Coop G., Przeworski M.. 2007, An evolutionary view of human recombination, Nat. Rev. Genet., 8, 23–34. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
