

Dependency of Heterochromatin Domains on Replication Factors
- PMID: 29187422
- PMCID: PMC5919735
- DOI: 10.1534/g3.117.300341
Dependency of Heterochromatin Domains on Replication Factors
Abstract
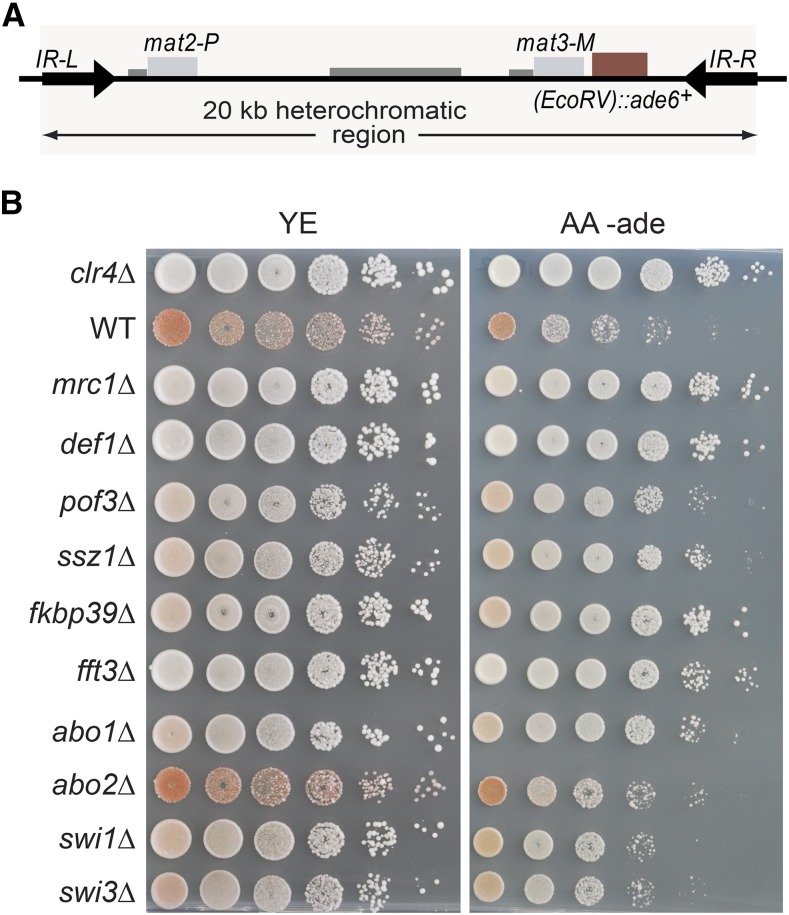
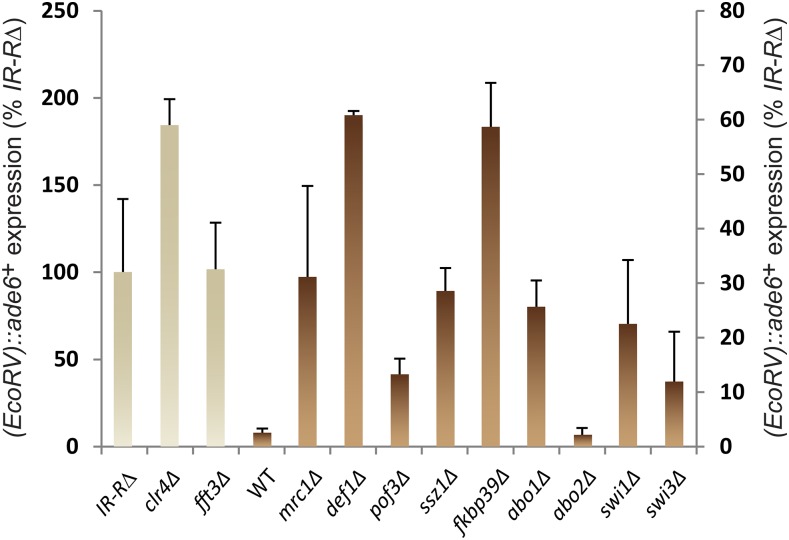
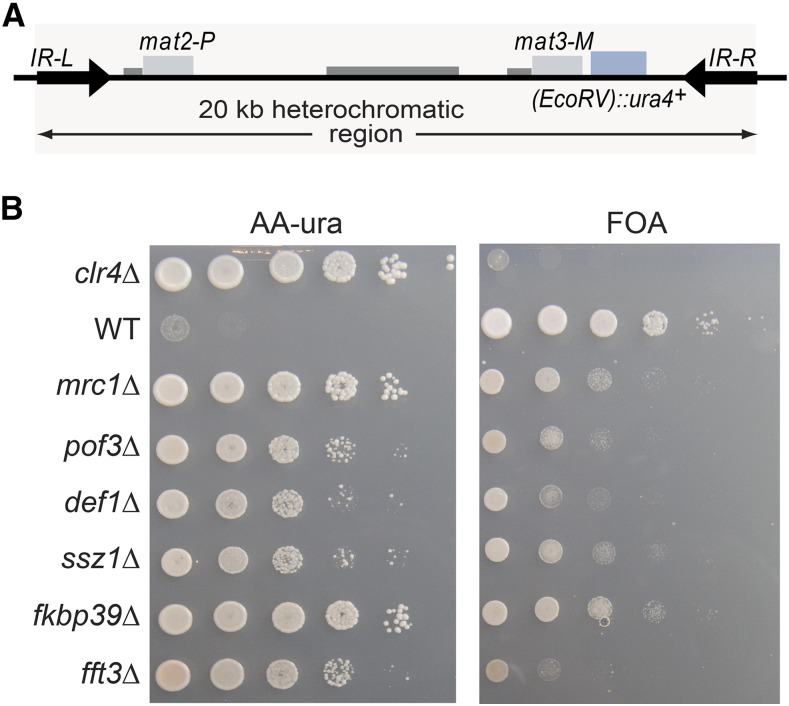
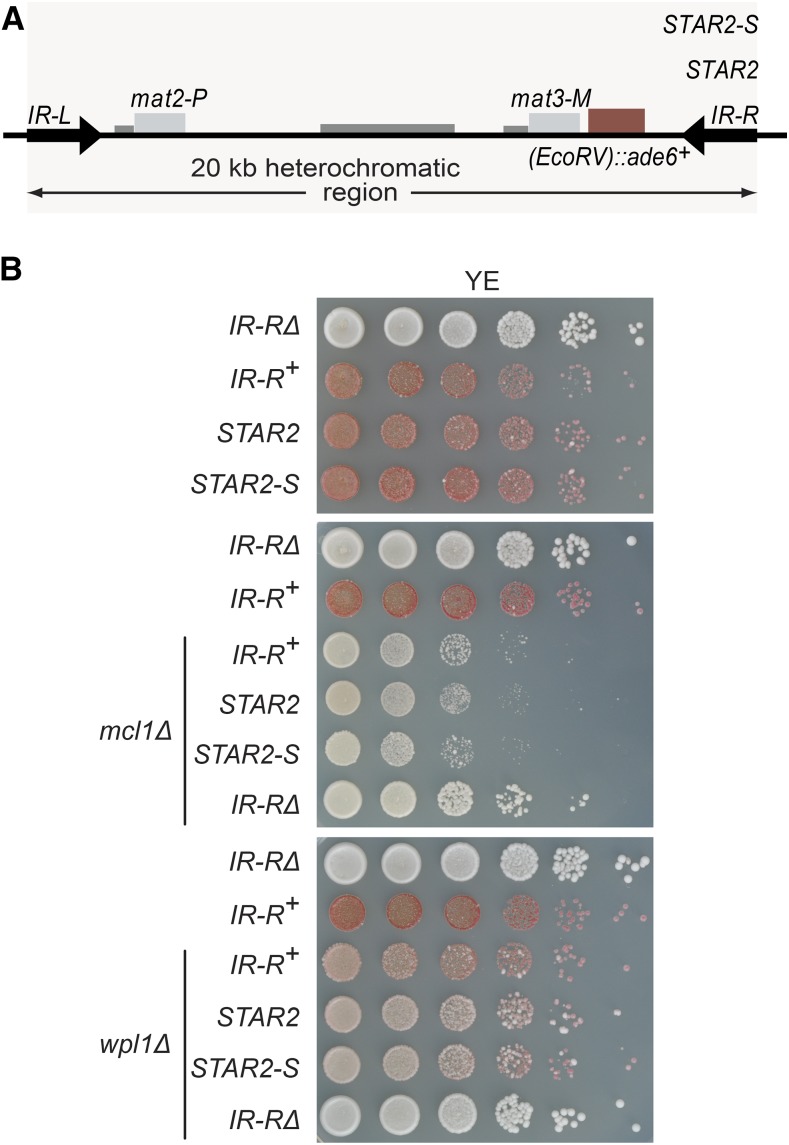
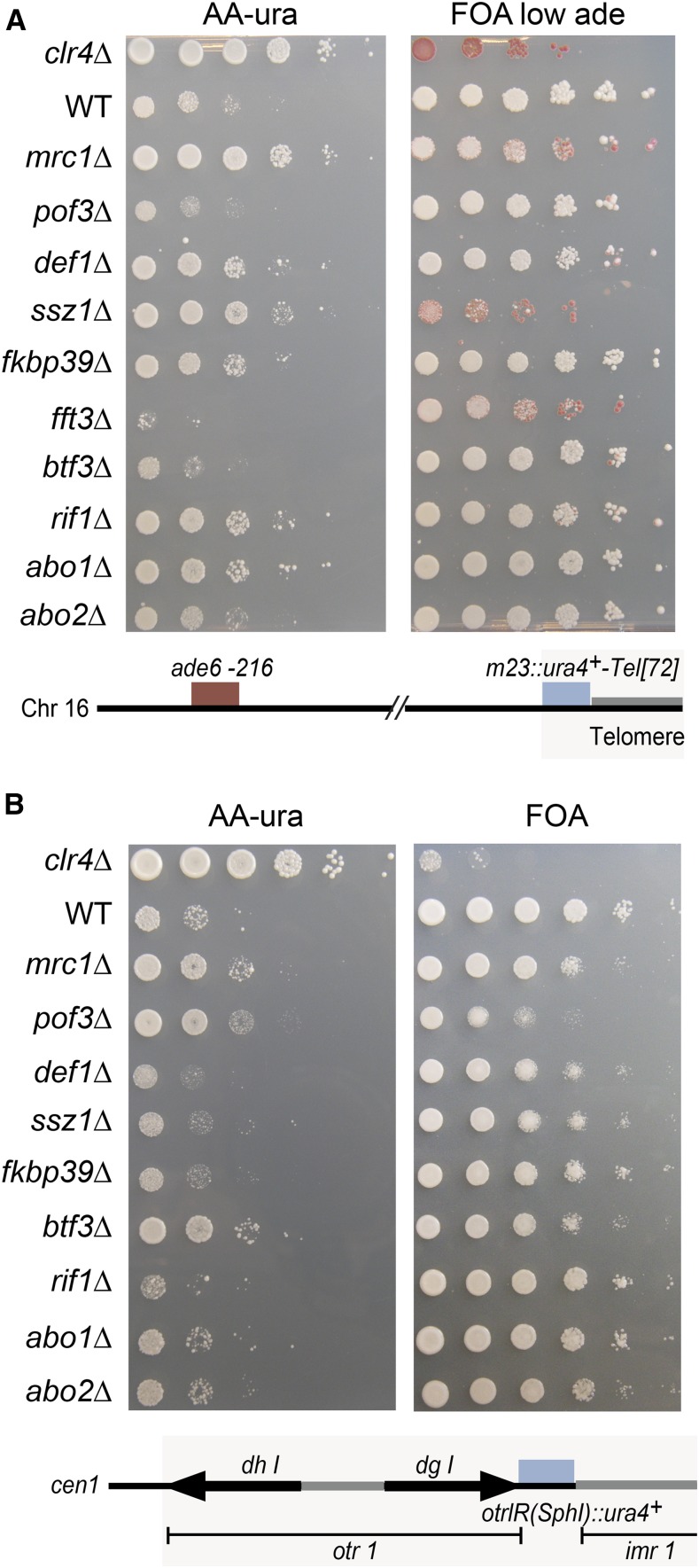
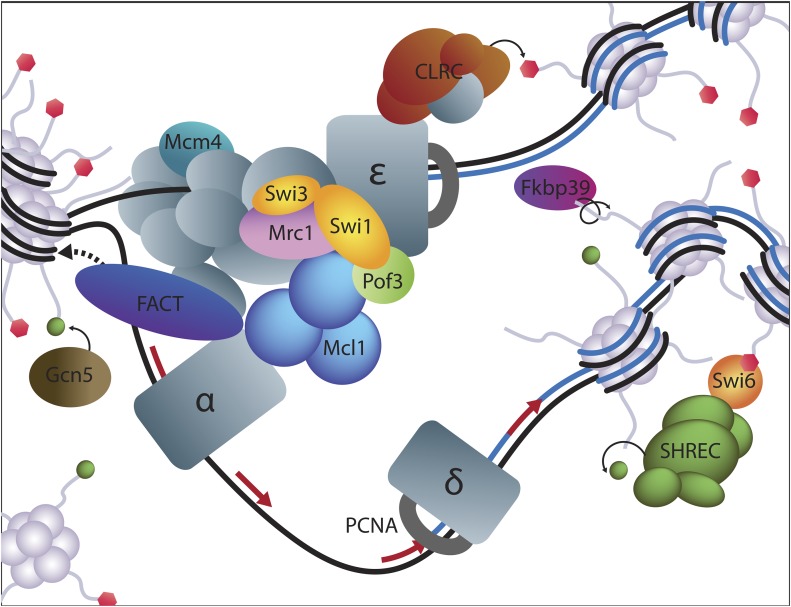
Chromatin structure regulates both genome expression and dynamics in eukaryotes, where large heterochromatic regions are epigenetically silenced through the methylation of histone H3K9, histone deacetylation, and the assembly of repressive complexes. Previous genetic screens with the fission yeast Schizosaccharomyces pombe have led to the identification of key enzymatic activities and structural constituents of heterochromatin. We report here on additional factors discovered by screening a library of deletion mutants for silencing defects at the edge of a heterochromatic domain bound by its natural boundary-the IR-R+ element-or by ectopic boundaries. We found that several components of the DNA replication progression complex (RPC), including Mrc1/Claspin, Mcl1/Ctf4, Swi1/Timeless, Swi3/Tipin, and the FACT subunit Pob3, are essential for robust heterochromatic silencing, as are the ubiquitin ligase components Pof3 and Def1, which have been implicated in the removal of stalled DNA and RNA polymerases from chromatin. Moreover, the search identified the cohesin release factor Wpl1 and the forkhead protein Fkh2, both likely to function through genome organization, the Ssz1 chaperone, the Fkbp39 proline cis-trans isomerase, which acts on histone H3P30 and P38 in Saccharomyces cerevisiae, and the chromatin remodeler Fft3. In addition to their effects in the mating-type region, to varying extents, these factors take part in heterochromatic silencing in pericentromeric regions and telomeres, revealing for many a general effect in heterochromatin. This list of factors provides precious new clues with which to study the spatiotemporal organization and dynamics of heterochromatic regions in connection with DNA replication.
Keywords: DNA replication; chromatin remodeling; cohesins; fission yeast; gene silencing; genetic screen; heterochromatin; mating-type region; nuclear organization; proline isomerase.
Copyright © 2018 Jahn et al.
Figures

References
-
- Allshire R. C., Nimmo E. R., Ekwall K., Javerzat J. P., Cranston G., 1995. Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes Dev. 9: 218–233. - PubMed
-
- Ansbach A. B., Noguchi C., Klansek I. W., Heidlebaugh M., Nakamura T. M., et al. , 2008. RFCCtf18 and the Swi1-Swi3 complex function in separate and redundant pathways required for the stabilization of replication forks to facilitate sister chromatid cohesion in Schizosaccharomyces pombe. Mol. Biol. Cell 19: 595–607. - PMC - PubMed
-
- Baryshnikova A., Costanzo M., Dixon S., Vizeacoumar F. J., Myers C. L., et al. , 2010. Synthetic genetic array (SGA) analysis in Saccharomyces cerevisiae and Schizosaccharomyces pombe. Methods Enzymol. 470: 145–179. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous