

To Be or Not To Be T4: Evidence of a Complex Evolutionary Pathway of Head Structure and Assembly in Giant Salmonella Virus SPN3US
- PMID: 29187846
- PMCID: PMC5694885
- DOI: 10.3389/fmicb.2017.02251
To Be or Not To Be T4: Evidence of a Complex Evolutionary Pathway of Head Structure and Assembly in Giant Salmonella Virus SPN3US
Abstract
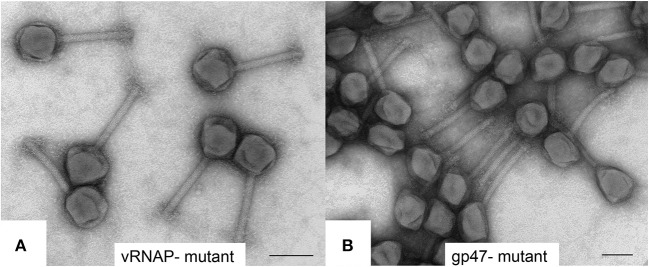
Giant Salmonella phage SPN3US has a 240-kb dsDNA genome and a large complex virion composed of many proteins for which the functions of most are undefined. We recently determined that SPN3US shares a core set of genes with related giant phages and sequenced and characterized 18 amber mutants to facilitate its use as a genetic model system. Notably, SPN3US and related giant phages contain a bolus of ejection proteins within their heads, including a multi-subunit virion RNA polymerase (vRNAP), that enter the host cell with the DNA during infection. In this study, we characterized the SPN3US virion using mass spectrometry to gain insight into its head composition and the features that its head shares with those of related giant phages and with T4 phage. SPN3US has only homologs to the T4 proteins critical for prohead shell formation, the portal and major capsid proteins, as well as to the major enzymes essential for head maturation, the prohead protease and large terminase subunit. Eight of ~50 SPN3US head proteins were found to undergo proteolytic processing at a cleavage motif by the prohead protease gp245. Gp245 undergoes auto-cleavage of its C-terminus, suggesting this is a conserved activation and/or maturation feature of related phage proteases. Analyses of essential head gene mutants showed that the five subunits of the vRNAP must be assembled for any subunit to be incorporated into the prohead, although the assembled vRNAP must then undergo subsequent major conformational rearrangements in the DNA packed capsid to allow ejection through the ~30 Å diameter tail tube for transcription from the injected DNA. In addition, ejection protein candidate gp243 was found to play a critical role in head assembly. Our analyses of the vRNAP and gp243 mutants highlighted an unexpected dichotomy in giant phage head maturation: while all analyzed giant phages have a homologous protease that processes major capsid and portal proteins, processing of ejection proteins is not always a stable/defining feature. Our identification in SPN3US, and related phages, of a diverged paralog to the prohead protease further hints toward a complicated evolutionary pathway for giant phage head structure and assembly.
Keywords: CTS (capsid targeting sequence); Salmonella; ejection protein; giant phage; mass spectrometry; myovirus; prohead protease; virion RNA polymerase (vRNAP).
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures









Similar articles
-
Global Proteomic Profiling of Salmonella Infection by a Giant Phage.J Virol. 2019 Feb 19;93(5):e01833-18. doi: 10.1128/JVI.01833-18. Print 2019 Mar 1. J Virol. 2019. PMID: 30541839 Free PMC article.
-
Identification of Essential Genes in the Salmonella Phage SPN3US Reveals Novel Insights into Giant Phage Head Structure and Assembly.J Virol. 2016 Oct 28;90(22):10284-10298. doi: 10.1128/JVI.01492-16. Print 2016 Nov 15. J Virol. 2016. PMID: 27605673 Free PMC article.
-
Mass Spectral Analyses of Salmonella Myovirus SPN3US Reveal Conserved and Divergent Themes in Proteolytic Maturation of Large Icosahedral Capsids.Viruses. 2023 Mar 10;15(3):723. doi: 10.3390/v15030723. Viruses. 2023. PMID: 36992431 Free PMC article.
-
Tailed bacteriophages: the order caudovirales.Adv Virus Res. 1998;51:135-201. doi: 10.1016/s0065-3527(08)60785-x. Adv Virus Res. 1998. PMID: 9891587 Free PMC article. Review.
-
Synthesis of a trans-acting inhibitor of DNA maturation by prohead mutants of phage lambda.Genetics. 1987 Jan;115(1):3-10. doi: 10.1093/genetics/115.1.3. Genetics. 1987. PMID: 2951296 Free PMC article. Review.
Cited by
-
Global Proteomic Profiling of Salmonella Infection by a Giant Phage.J Virol. 2019 Feb 19;93(5):e01833-18. doi: 10.1128/JVI.01833-18. Print 2019 Mar 1. J Virol. 2019. PMID: 30541839 Free PMC article.
-
Genomic Characterization of Jumbo Salmonella Phages That Effectively Target United Kingdom Pig-Associated Salmonella Serotypes.Front Microbiol. 2019 Jul 2;10:1491. doi: 10.3389/fmicb.2019.01491. eCollection 2019. Front Microbiol. 2019. PMID: 31312191 Free PMC article.
-
Intravirion DNA Can Access the Space Occupied by the Bacteriophage P22 Ejection Proteins.Viruses. 2021 Jul 30;13(8):1504. doi: 10.3390/v13081504. Viruses. 2021. PMID: 34452369 Free PMC article.
-
Isolation and Characterization of Salmonella Jumbo-Phage pSal-SNUABM-04.Viruses. 2020 Dec 25;13(1):27. doi: 10.3390/v13010027. Viruses. 2020. PMID: 33375688 Free PMC article.
-
A Cut above the Rest: Characterization of the Assembly of a Large Viral Icosahedral Capsid.Viruses. 2020 Jul 5;12(7):725. doi: 10.3390/v12070725. Viruses. 2020. PMID: 32635654 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials