

Responses to a warming world: Integrating life history, immune investment, and pathogen resistance in a model insect species
- PMID: 29188001
- PMCID: PMC5696387
- DOI: 10.1002/ece3.3506
Responses to a warming world: Integrating life history, immune investment, and pathogen resistance in a model insect species
Abstract
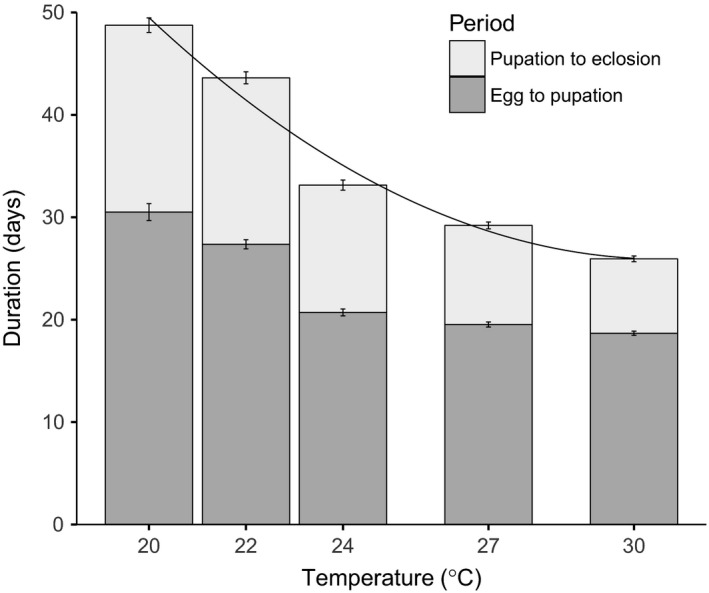
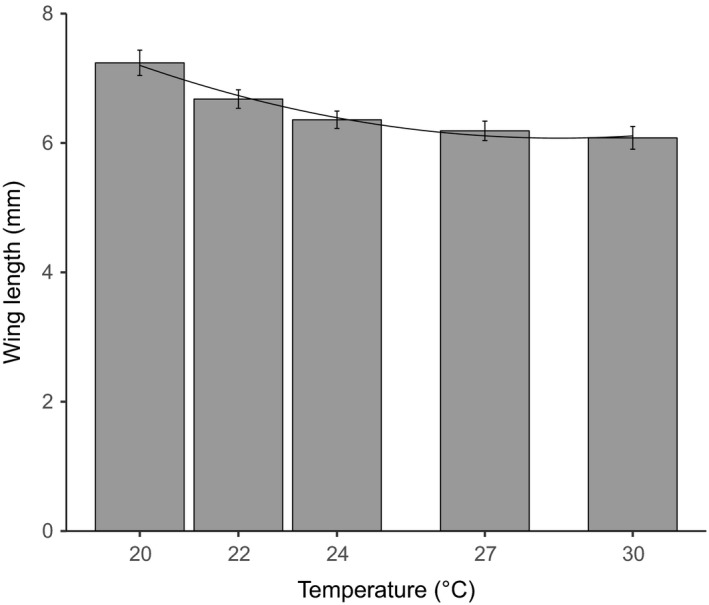
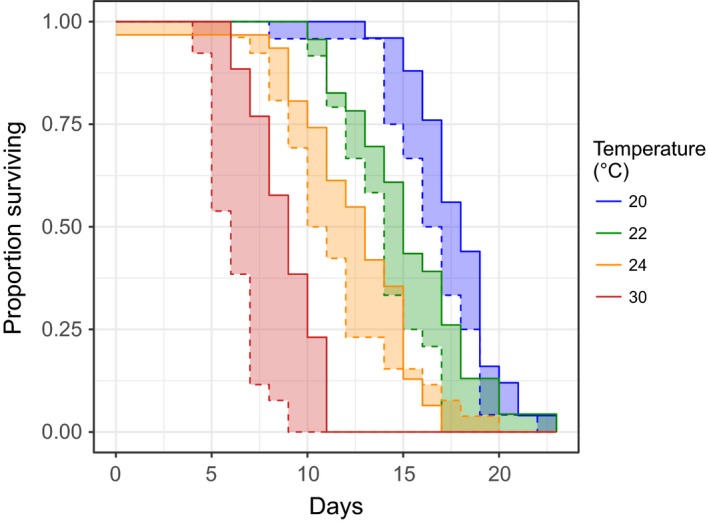
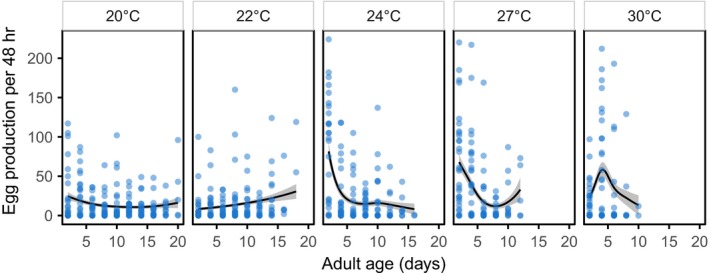
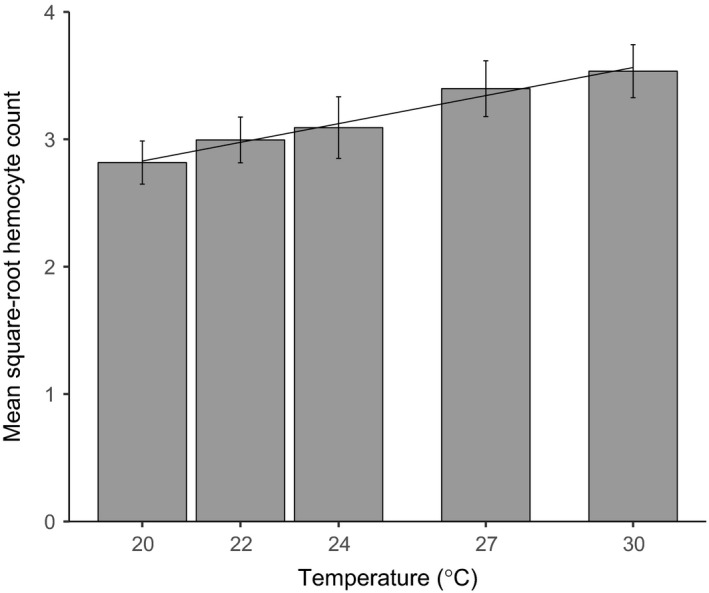
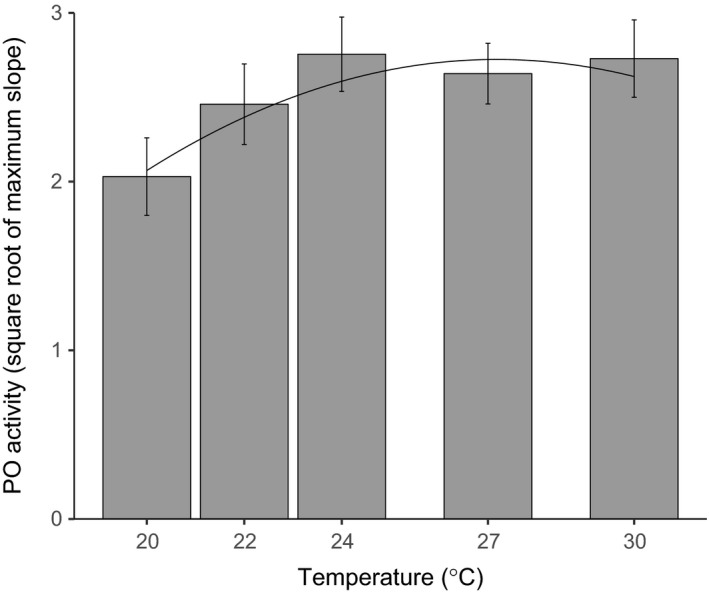
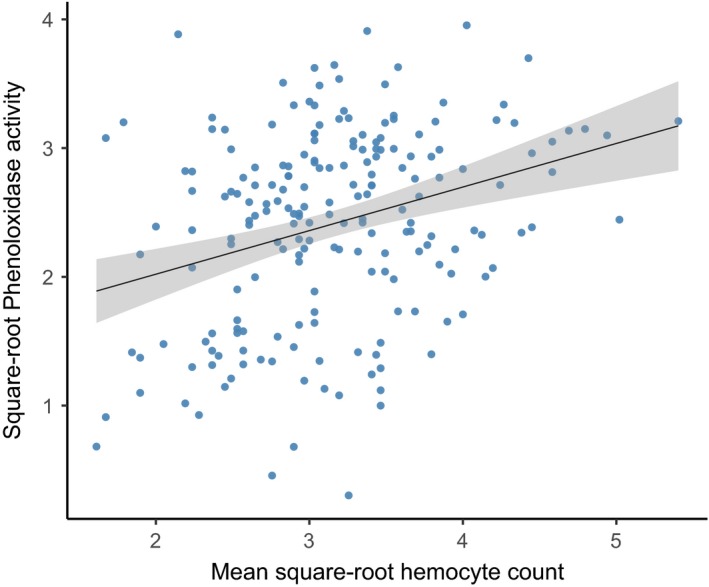
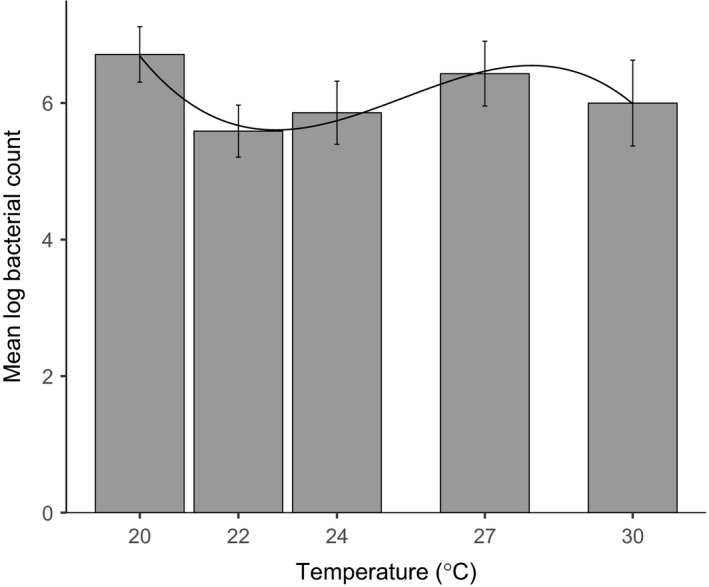
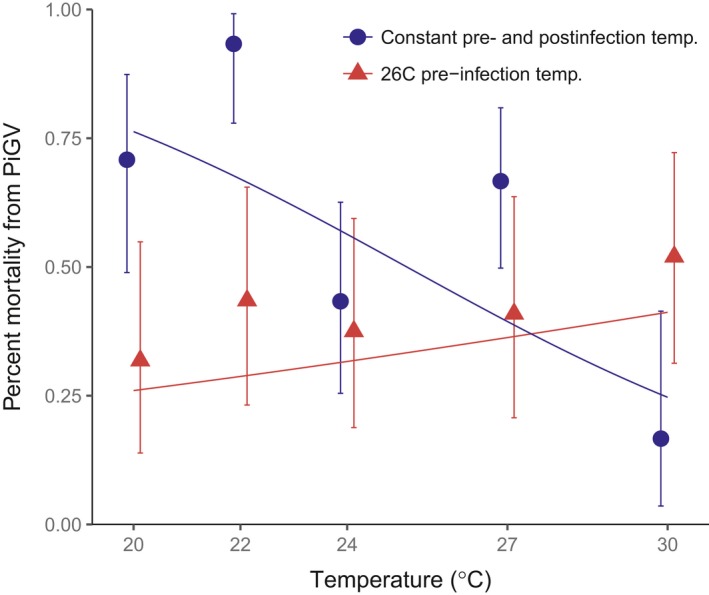
Environmental temperature has important effects on the physiology and life history of ectothermic animals, including investment in the immune system and the infectious capacity of pathogens. Numerous studies have examined individual components of these complex systems, but little is known about how they integrate when animals are exposed to different temperatures. Here, we use the Indian meal moth (Plodia interpunctella) to understand how immune investment and disease resistance react and potentially trade-off with other life-history traits. We recorded life-history (development time, survival, fecundity, and body size) and immunity (hemocyte counts, phenoloxidase activity) measures and tested resistance to bacterial (E. coli) and viral (Plodia interpunctella granulosis virus) infection at five temperatures (20-30°C). While development time, lifespan, and size decreased with temperature as expected, moths exhibited different reproductive strategies in response to small changes in temperature. At cooler temperatures, oviposition rates were low but tended to increase toward the end of life, whereas warmer temperatures promoted initially high oviposition rates that rapidly declined after the first few days of adult life. Although warmer temperatures were associated with strong investment in early reproduction, there was no evidence of an associated trade-off with immune investment. Phenoloxidase activity increased most at cooler temperatures before plateauing, while hemocyte counts increased linearly with temperature. Resistance to bacterial challenge displayed a complex pattern, whereas survival after a viral challenge increased with rearing temperature. These results demonstrate that different immune system components and different pathogens can respond in distinct ways to changes in temperature. Overall, these data highlight the scope for significant changes in immunity, disease resistance, and host-parasite population dynamics to arise from small, biologically relevant changes to environmental temperature. In light of global warming, understanding these complex interactions is vital for predicting the potential impact of insect disease vectors and crop pests on public health and food security.
Keywords: Plodia interpunctella; defense; ecological immunology; global warming; hemocyte; phenoloxidase; trade‐off.
Figures
Similar articles
-
Interactions between environmental variables determine immunity in the Indian meal moth Plodia interpunctella.J Anim Ecol. 2012 Mar;81(2):386-94. doi: 10.1111/j.1365-2656.2011.01920.x. Epub 2011 Oct 14. J Anim Ecol. 2012. PMID: 21999965
-
Consequences of Food Restriction for Immune Defense, Parasite Infection, and Fitness in Monarch Butterflies.Physiol Biochem Zool. 2016 Sep-Oct;89(5):389-401. doi: 10.1086/687989. Epub 2016 Jul 25. Physiol Biochem Zool. 2016. PMID: 27617360
-
Experimental evolution reveals differences between phenotypic and evolutionary responses to population density.J Evol Biol. 2017 Sep;30(9):1763-1771. doi: 10.1111/jeb.13139. Epub 2017 Aug 4. J Evol Biol. 2017. PMID: 28675768
-
The role of fecundity and reproductive effort in defining life-history strategies of North American freshwater mussels.Biol Rev Camb Philos Soc. 2013 Aug;88(3):745-66. doi: 10.1111/brv.12028. Epub 2013 Feb 28. Biol Rev Camb Philos Soc. 2013. PMID: 23445204 Review.
-
Granulosis viruses, with emphasis on the GV of the Indian meal moth, Plodia interpunctella.Adv Virus Res. 1983;28:141-73. doi: 10.1016/s0065-3527(08)60723-x. Adv Virus Res. 1983. PMID: 6362364 Review.
Cited by
-
Warming at the population level: Effects on age structure, density, and generation cycles.Ecol Evol. 2019 Mar 28;9(8):4403-4420. doi: 10.1002/ece3.4972. eCollection 2019 Apr. Ecol Evol. 2019. PMID: 31031915 Free PMC article.
-
Comparative transcriptome analysis of the rice leaf folder (Cnaphalocrocis medinalis) to heat acclimation.BMC Genomics. 2020 Jun 30;21(1):450. doi: 10.1186/s12864-020-06867-6. BMC Genomics. 2020. PMID: 32605538 Free PMC article.
-
Immune Defenses of a Beneficial Pest: The Mealworm Beetle, Tenebrio molitor.Front Physiol. 2019 Mar 12;10:138. doi: 10.3389/fphys.2019.00138. eCollection 2019. Front Physiol. 2019. PMID: 30914960 Free PMC article. Review.
-
Developmental Temperature Affects Life-History Traits and Heat Tolerance in the Aphid Parasitoid Aphidius colemani.Insects. 2021 Sep 22;12(10):852. doi: 10.3390/insects12100852. Insects. 2021. PMID: 34680621 Free PMC article.
-
Impact of the Diet on the Mortality and on Gene Expression of the Antimicrobial Peptide Tenecin 3 in Tenebrio molitor Larvae Infected by Beauveria bassiana.Insects. 2023 Apr 4;14(4):359. doi: 10.3390/insects14040359. Insects. 2023. PMID: 37103174 Free PMC article.
References
-
- Adamo, S. A. , & Lovett, M. M. E. (2011). Some like it hot: The effects of climate change on reproduction, immune function and disease resistance in the cricket Gryllus texensis. The Journal of Experimental Biology, 214, 1997–2004. - PubMed
-
- Aguilar‐Alberola, J. A. , & Mesquita‐Joanes, F. (2014). Breaking the temperature‐size rule: Thermal effects on growth, development and fecundity of a crustacean from temporary waters. Journal of Thermal Biology, 42, 15–24. - PubMed
-
- Angilletta, M. J. , & Dunham, A. E. (2003). The temperature‐size rule in ectotherms: Simple evolutionary explanations may not be general. American Naturalist, 162, 332–342. - PubMed
-
- Angilletta, M. J. , Huey, R. B. , & Frazier, M. R. (2010). Thermodynamic effects on organismal performance: Is hotter better? Physiological and Biochemical Zoology, 83, 197–206. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
