

Radiation of the polymorphic Little Devil poison frog (Oophaga sylvatica) in Ecuador
- PMID: 29188006
- PMCID: PMC5696431
- DOI: 10.1002/ece3.3503
Radiation of the polymorphic Little Devil poison frog (Oophaga sylvatica) in Ecuador
Abstract
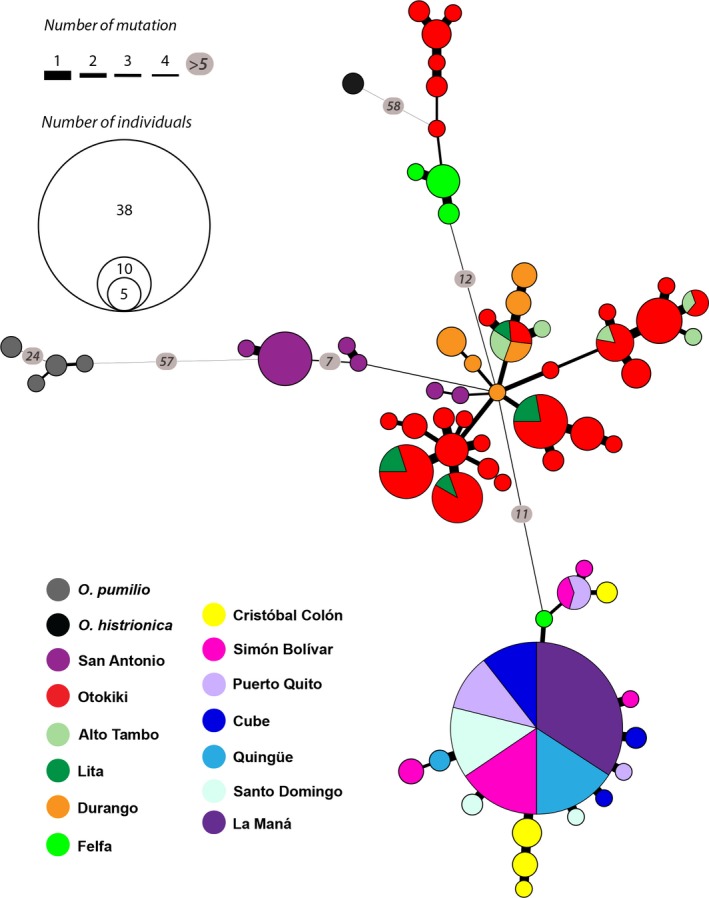
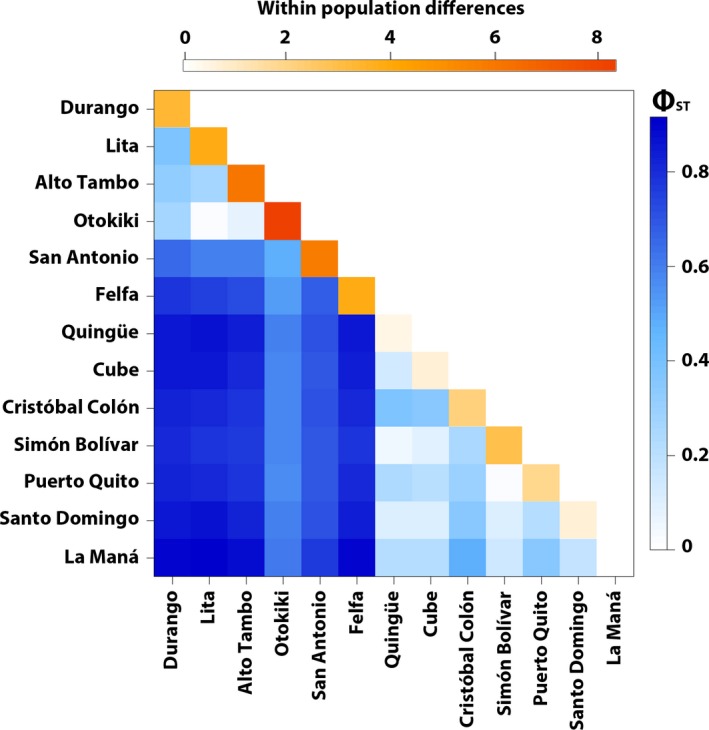
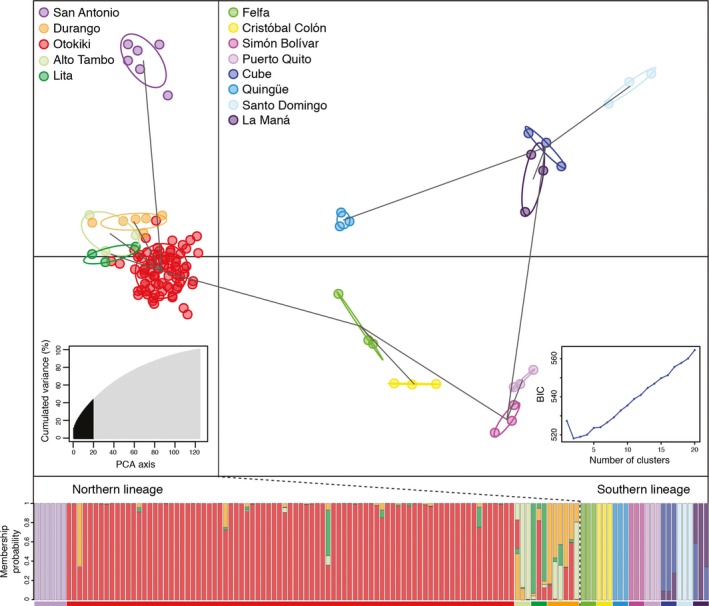
Some South American poison frogs (Dendrobatidae) are chemically defended and use bright aposematic colors to warn potential predators of their unpalatability. Aposematic signals are often frequency-dependent where individuals deviating from a local model are at a higher risk of predation. However, extreme diversity in the aposematic signal has been documented in poison frogs, especially in Oophaga. Here, we explore the phylogeographic pattern among color-divergent populations of the Little Devil poison frog Oophaga sylvatica by analyzing population structure and genetic differentiation to evaluate which processes could account for color diversity within and among populations. With a combination of PCR amplicons (three mitochondrial and three nuclear markers) and genome-wide markers from a double-digested RAD (ddRAD) approach, we characterized the phylogenetic and genetic structure of 199 individuals from 13 populations (12 monomorphic and 1 polymorphic) across the O. sylvatica distribution. Individuals segregated into two main lineages by their northern or southern latitudinal distribution. A high level of genetic and phenotypic polymorphism within the northern lineage suggests ongoing gene flow. In contrast, low levels of genetic differentiation were detected among the southern lineage populations and support recent range expansions from populations in the northern lineage. We propose that a combination of climatic gradients and structured landscapes might be promoting gene flow and phylogenetic diversification. Alternatively, we cannot rule out that the observed phenotypic and genomic variations are the result of genetic drift on near or neutral alleles in a small number of genes.
Keywords: Dendrobatidae; Ecuador; Oophaga sylvatica; amphibian; aposematism; ddRAD; gene flow; phenotypic variation; population genomics.
Figures



References
-
- Arteaga, A. , Pyron, R. A. , Peñafiel, N. , Romero‐Barreto, P. , Culebras, J. , Bustamante, L. , … Guayasamin, J. M. (2016). Comparative phylogeography reveals cryptic diversity and repeated patterns of cladogenesis for amphibians and reptiles in northwestern Ecuador. PLoS ONE, 11, e0151746. - PMC - PubMed
-
- Bastian, M. , Heymann, S. , & Jacomy, M. (2009). Gephi: an open source software for exploring and manipulating networks. ICWSM, 8, 361–362.
-
- Benson, W. W. (1971). Evidence for the evolution of unpalatability through kin selection in the Heliconiinae (Lepidoptera). The American Naturalist, 105, 213.
-
- Brodie, E. D. I. (1993). Differential avoidance of coral snake banded patterns by free‐ranging avian predators in Costa Rica. Evolution, 47, 227–235. - PubMed
-
- Brown, J. , Twomey, E. , Amézquita, A. , de Souza, M. B. , Caldwell, J. P. , Lötters, S. , … Summers, K. (2011). A taxonomic revision of the Neotropical poison frog genus Ranitomeya (Amphibia: Dendrobatidae). Zootaxa, 120, 1–120.
LinkOut - more resources
Full Text Sources
Other Literature Sources
