

NudE regulates dynein at kinetochores but is dispensable for other dynein functions in the C. elegans early embryo
- PMID: 29192061
- PMCID: PMC5818066
- DOI: 10.1242/jcs.212159
NudE regulates dynein at kinetochores but is dispensable for other dynein functions in the C. elegans early embryo
Abstract
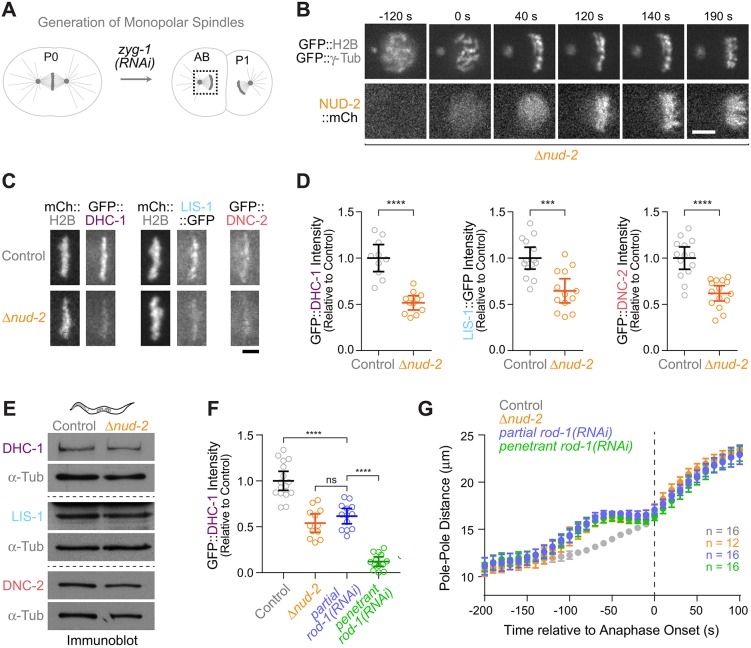
In mitosis, the molecular motor dynein is recruited to kinetochores by the Rod-Zw10-Zwilch complex (RZZ) and Spindly to control spindle assembly checkpoint (SAC) signaling and microtubule attachment. How the ubiquitous dynein co-factors Lis1 and NudE contribute to these functions remains poorly understood. Here, we show that the C. elegans NudE homolog NUD-2 is dispensable for dynein- and LIS-1-dependent mitotic spindle assembly in the zygote. This facilitates functional characterization of kinetochore-localized NUD-2, which is recruited by the CENP-F-like proteins HCP-1 and HCP-2 independently of RZZ-Spindly and dynein-LIS-1. Kinetochore dynein levels are reduced in Δnud-2 embryos, and, as occurs upon RZZ inhibition, loss of NUD-2 delays the formation of load-bearing kinetochore-microtubule attachments and causes chromatin bridges in anaphase. Survival of Δnud-2 embryos requires a functional SAC, and kinetochores without NUD-2 recruit an excess of SAC proteins. Consistent with this, SAC signaling in early Δnud-2 embryos extends mitotic duration and prevents high rates of chromosome mis-segregation. Our results reveal that both NUD-2 and RZZ-Spindly are essential for dynein function at kinetochores, and that the gain in SAC strength during early embryonic development is relevant under conditions that mildly perturb mitosis.
Keywords: Dynein; Kinetochore; Lis1; NDE1; NudE; PAFAH1B1; RZZ; Spindle assembly checkpoint.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures





Similar articles
-
Molecular mechanism of dynein recruitment to kinetochores by the Rod-Zw10-Zwilch complex and Spindly.J Cell Biol. 2017 Apr 3;216(4):943-960. doi: 10.1083/jcb.201610108. Epub 2017 Mar 20. J Cell Biol. 2017. PMID: 28320824 Free PMC article.
-
Self-Assembly of the RZZ Complex into Filaments Drives Kinetochore Expansion in the Absence of Microtubule Attachment.Curr Biol. 2018 Nov 5;28(21):3408-3421.e8. doi: 10.1016/j.cub.2018.08.056. Epub 2018 Oct 25. Curr Biol. 2018. PMID: 30415699 Free PMC article.
-
A new mechanism controlling kinetochore-microtubule interactions revealed by comparison of two dynein-targeting components: SPDL-1 and the Rod/Zwilch/Zw10 complex.Genes Dev. 2008 Sep 1;22(17):2385-99. doi: 10.1101/gad.1687508. Genes Dev. 2008. PMID: 18765790 Free PMC article.
-
Spindly switch controls anaphase: spindly and RZZ functions in chromosome attachment and mitotic checkpoint control.Cell Cycle. 2011 Feb 1;10(3):449-56. doi: 10.4161/cc.10.3.14759. Epub 2011 Feb 1. Cell Cycle. 2011. PMID: 21252629 Review.
-
RZZ-SPINDLY-DYNEIN: you got to keep 'em separated.Cell Cycle. 2020 Jul;19(14):1716-1726. doi: 10.1080/15384101.2020.1780382. Epub 2020 Jun 16. Cell Cycle. 2020. PMID: 32544383 Free PMC article. Review.
Cited by
-
Synergistic stabilization of microtubules by BUB-1, HCP-1, and CLS-2 controls microtubule pausing and meiotic spindle assembly.Elife. 2023 Feb 17;12:e82579. doi: 10.7554/eLife.82579. Elife. 2023. PMID: 36799894 Free PMC article.
-
The kinetochore proteins CENP-E and CENP-F directly and specifically interact with distinct BUB mitotic checkpoint Ser/Thr kinases.J Biol Chem. 2018 Jun 29;293(26):10084-10101. doi: 10.1074/jbc.RA118.003154. Epub 2018 May 10. J Biol Chem. 2018. PMID: 29748388 Free PMC article.
-
Cargo-Mediated Activation of Cytoplasmic Dynein in vivo.Front Cell Dev Biol. 2020 Oct 23;8:598952. doi: 10.3389/fcell.2020.598952. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33195284 Free PMC article. Review.
-
A transient helix in the disordered region of dynein light intermediate chain links the motor to structurally diverse adaptors for cargo transport.PLoS Biol. 2019 Jan 7;17(1):e3000100. doi: 10.1371/journal.pbio.3000100. eCollection 2019 Jan. PLoS Biol. 2019. PMID: 30615611 Free PMC article.
-
Lissencephaly-1 mutations enhance traumatic brain injury outcomes in Drosophila.Genetics. 2023 Mar 2;223(3):iyad008. doi: 10.1093/genetics/iyad008. Genetics. 2023. PMID: 36683334 Free PMC article.
References
-
- Barbosa D. J., Duro J., Prevo B., Cheerambathur D. K., Carvalho A. X. and Gassmann R. (2017). Dynactin binding to tyrosinated microtubules promotes centrosome centration in C. elegans by enhancing dynein-mediated organelle transport. PLoS Genet. 13, e1006941 10.1371/journal.pgen.1006941 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
