

Developmental YAPdeltaC determines adult pathology in a model of spinocerebellar ataxia type 1
- PMID: 29192206
- PMCID: PMC5709507
- DOI: 10.1038/s41467-017-01790-z
Developmental YAPdeltaC determines adult pathology in a model of spinocerebellar ataxia type 1
Abstract
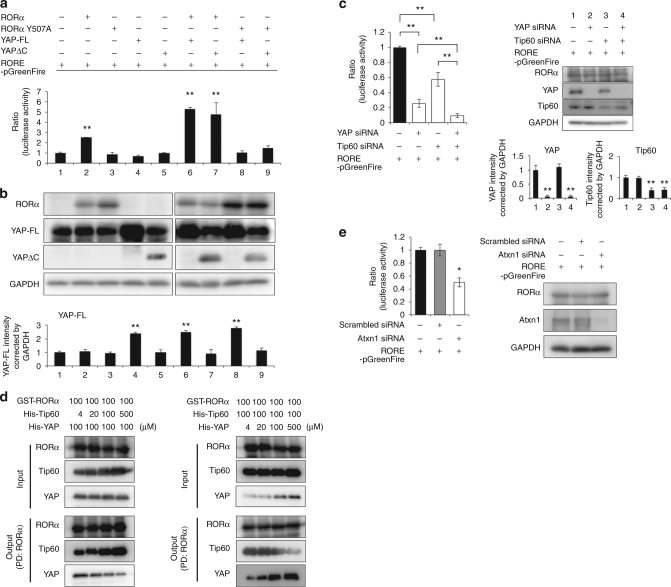
YAP and its neuronal isoform YAPdeltaC are implicated in various cellular functions. We found that expression of YAPdeltaC during development, but not adulthood, rescued neurodegeneration phenotypes of mutant ataxin-1 knock-in (Atxn1-KI) mice. YAP/YAPdeltaC interacted with RORα via the second WW domain and served as co-activators of its transcriptional activity. YAP/YAPdeltaC formed a transcriptional complex with RORα on cis-elements of target genes and regulated their expression. Both normal and mutant Atxn1 interacted with YAP/YAPdeltaC, but only mutant Atxn1 depleted YAP/YAPdeltaC from the RORα complex to suppress transcription on short timescales. Over longer periods, mutant Atxn1 also decreased RORα in vivo. Genetic supplementation of YAPdeltaC restored the RORα and YAP/YAPdeltaC levels, recovered YAP/YAPdeltaC in the RORα complex and normalized target gene transcription in Atxn1-KI mice in vivo. Collectively, our data suggest that functional impairment of YAP/YAPdeltaC by mutant Atxn1 during development determines the adult pathology of SCA1 by suppressing RORα-mediated transcription.
Conflict of interest statement
The authors declare no competing financial interests.
Figures









Similar articles
-
Ataxin-1 regulates the cerebellar bioenergetics proteome through the GSK3β-mTOR pathway which is altered in Spinocerebellar ataxia type 1 (SCA1).Hum Mol Genet. 2016 Sep 15;25(18):4021-4040. doi: 10.1093/hmg/ddw242. Epub 2016 Jul 27. Hum Mol Genet. 2016. PMID: 27466200
-
Partial loss of Tip60 slows mid-stage neurodegeneration in a spinocerebellar ataxia type 1 (SCA1) mouse model.Hum Mol Genet. 2011 Jun 1;20(11):2204-12. doi: 10.1093/hmg/ddr108. Epub 2011 Mar 22. Hum Mol Genet. 2011. PMID: 21427130 Free PMC article.
-
Indirect Negative Effect of Mutant Ataxin-1 on Short- and Long-Term Synaptic Plasticity in Mouse Models of Spinocerebellar Ataxia Type 1.Cells. 2022 Jul 20;11(14):2247. doi: 10.3390/cells11142247. Cells. 2022. PMID: 35883691 Free PMC article.
-
UTteR control through miRs: fine-tuning ATXN1 levels to prevent ataxia.Genes Dev. 2020 Sep 1;34(17-18):1107-1109. doi: 10.1101/gad.343020.120. Genes Dev. 2020. PMID: 32873576 Free PMC article. Review.
-
Spinocerebellar Ataxia Type 1: Molecular Mechanisms of Neurodegeneration and Preclinical Studies.Adv Exp Med Biol. 2018;1049:135-145. doi: 10.1007/978-3-319-71779-1_6. Adv Exp Med Biol. 2018. PMID: 29427101 Review.
Cited by
-
HD and SCA1: Tales from two 30-year journeys since gene discovery.Neuron. 2023 Nov 15;111(22):3517-3530. doi: 10.1016/j.neuron.2023.09.036. Epub 2023 Oct 19. Neuron. 2023. PMID: 37863037 Free PMC article. Review.
-
Dynamic molecular network analysis of iPSC-Purkinje cells differentiation delineates roles of ISG15 in SCA1 at the earliest stage.Commun Biol. 2024 Apr 9;7(1):413. doi: 10.1038/s42003-024-06066-z. Commun Biol. 2024. PMID: 38594382 Free PMC article.
-
Identification of the ataxin-1 interaction network and its impact on spinocerebellar ataxia type 1.Hum Genomics. 2022 Jul 29;16(1):29. doi: 10.1186/s40246-022-00404-0. Hum Genomics. 2022. PMID: 35906672 Free PMC article.
-
Dysfunctional Mechanotransduction through the YAP/TAZ/Hippo Pathway as a Feature of Chronic Disease.Cells. 2020 Jan 8;9(1):151. doi: 10.3390/cells9010151. Cells. 2020. PMID: 31936297 Free PMC article. Review.
-
Alternative Splicing in the Hippo Pathway-Implications for Disease and Potential Therapeutic Targets.Genes (Basel). 2018 Mar 13;9(3):161. doi: 10.3390/genes9030161. Genes (Basel). 2018. PMID: 29534050 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
