

Identification of a novel compound targeting the nuclear export of influenza A virus nucleoprotein
- PMID: 29193684
- PMCID: PMC5824420
- DOI: 10.1111/jcmm.13467
Identification of a novel compound targeting the nuclear export of influenza A virus nucleoprotein
Abstract
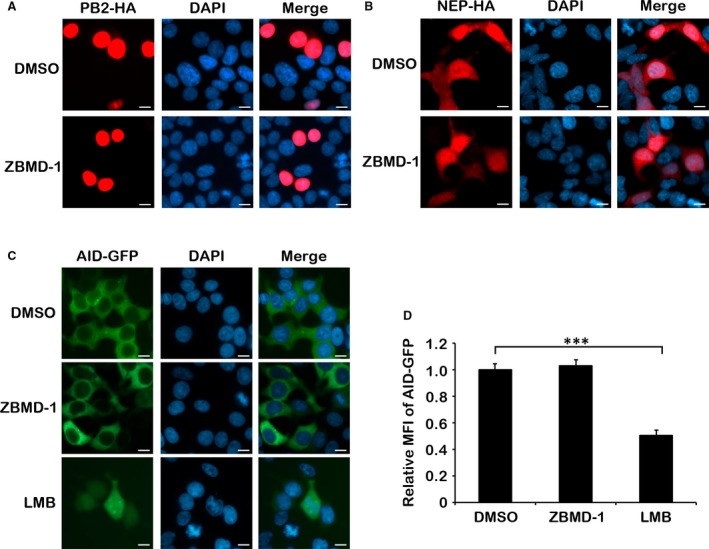
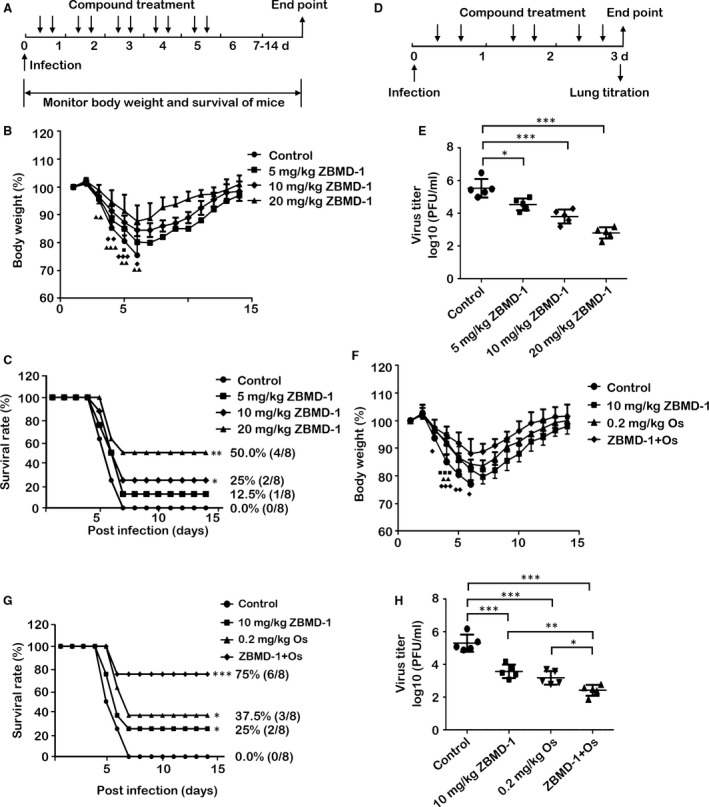
Although antiviral drugs are available for the treatment of influenza infection, it is an urgent requirement to develop new antiviral drugs regarding the emergence of drug-resistant viruses. The nucleoprotein (NP) is conserved among all influenza A viruses (IAVs) and has no cellular equivalent. Therefore, NP is an ideal target for the development of new IAV inhibitors. In this study, we identified a novel anti-influenza compound, ZBMD-1, from a library of 20,000 compounds using cell-based influenza A infection assays. We found that ZBMD-1 inhibited the replication of H1N1 and H3N2 influenza A virus strains in vitro, with an IC50 ranging from 0.41-1.14 μM. Furthermore, ZBMD-1 inhibited the polymerase activity and specifically impaired the nuclear export of NP. Further investigation indicated that ZBMD-1 binds to the nuclear export signal 3 (NES3) domain and the dimer interface of the NP pocket. ZBMD-1 also protected mice that were challenged with lethal doses of A/PR/8/1934 (H1N1) virus, effectively relieving lung histopathology changes, as well as strongly inhibiting the expression of pro-inflammatory cytokines/chemokines, without inducing toxicity effects in mice. These results suggest that ZBMD-1 is a promising anti-influenza compound which can be further investigated as a useful strategy against IAVs in the future.
Keywords: compound ZBMD-1; influenza A virus; nuclear export; nucleoprotein.
© 2017 The Authors. Journal of Cellular and Molecular Medicine published by John Wiley & Sons Ltd and Foundation for Cellular and Molecular Medicine.
Figures





Similar articles
-
A Novel Antiviral Target Structure Involved in the RNA Binding, Dimerization, and Nuclear Export Functions of the Influenza A Virus Nucleoprotein.PLoS Pathog. 2015 Jul 29;11(7):e1005062. doi: 10.1371/journal.ppat.1005062. eCollection 2015 Jul. PLoS Pathog. 2015. PMID: 26222066 Free PMC article.
-
A high-throughput screening system targeting the nuclear export pathway via the third nuclear export signal of influenza A virus nucleoprotein.Virus Res. 2016 Jun 2;217:23-31. doi: 10.1016/j.virusres.2016.02.007. Epub 2016 Mar 4. Virus Res. 2016. PMID: 26948263
-
Inhibition of influenza virus replication by constrained peptides targeting nucleoprotein.Antivir Chem Chemother. 2011 Nov 17;22(3):119-30. doi: 10.3851/IMP1902. Antivir Chem Chemother. 2011. PMID: 22095520
-
Influenza A Virus Nucleoprotein: A Highly Conserved Multi-Functional Viral Protein as a Hot Antiviral Drug Target.Curr Top Med Chem. 2017;17(20):2271-2285. doi: 10.2174/1568026617666170224122508. Curr Top Med Chem. 2017. PMID: 28240183 Free PMC article. Review.
-
Antiviral drug development by targeting RNA binding site, oligomerization and nuclear export of influenza nucleoprotein.Int J Biol Macromol. 2024 Dec;282(Pt 4):136996. doi: 10.1016/j.ijbiomac.2024.136996. Epub 2024 Oct 31. Int J Biol Macromol. 2024. PMID: 39486729 Review.
Cited by
-
Antiviral strategies against influenza virus: an update on approved and innovative therapeutic approaches.Cell Mol Life Sci. 2025 Feb 13;82(1):75. doi: 10.1007/s00018-025-05611-1. Cell Mol Life Sci. 2025. PMID: 39945883 Free PMC article. Review.
-
CD4+ TSCMs in the Bone Marrow Assist in Maturation of Antibodies against Influenza in Mice.Mediators Inflamm. 2019 Jan 10;2019:3231696. doi: 10.1155/2019/3231696. eCollection 2019. Mediators Inflamm. 2019. PMID: 30733641 Free PMC article.
-
Recent advances of phenotypic screening strategies in the application of anti-influenza virus drug discovery.RSC Med Chem. 2023 Nov 9;15(1):70-80. doi: 10.1039/d3md00513e. eCollection 2024 Jan 25. RSC Med Chem. 2023. PMID: 38283223 Free PMC article. Review.
-
Contemporary medicinal chemistry strategies for the discovery and optimization of influenza inhibitors targeting vRNP constituent proteins.Acta Pharm Sin B. 2022 Apr;12(4):1805-1824. doi: 10.1016/j.apsb.2021.11.018. Epub 2021 Nov 19. Acta Pharm Sin B. 2022. PMID: 35847499 Free PMC article. Review.
-
BRD7 as key factor in PBAF complex assembly and CD8+ T cell differentiation.JCI Insight. 2024 Jul 2;9(15):e171605. doi: 10.1172/jci.insight.171605. JCI Insight. 2024. PMID: 38954484 Free PMC article.
References
-
- Molinari NA, Ortega‐Sanchez IR, Messonnier ML, et al The annual impact of seasonal influenza in the US: measuring disease burden and costs. Vaccine. 2007; 25: 5086–96. - PubMed
-
- Simonsen L. The global impact of influenza on morbidity and mortality. Vaccine. 1999; 17(suppl 1): S3–10. - PubMed
-
- van der Vries E, Schutten M, Fraaij P, et al Influenza virus resistance to antiviral therapy. Adv Pharmacol. 2013; 67: 217–46. - PubMed
-
- Oxford JS, Mann A, Lambkin R. A designer drug against influenza: the NA inhibitor oseltamivir (Tamiflu). Expert Rev Anti Infect Ther. 2003; 1: 337–42. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
