

An Unexpectedly Complex Architecture for Skin Pigmentation in Africans
- PMID: 29195075
- PMCID: PMC5884124
- DOI: 10.1016/j.cell.2017.11.015
An Unexpectedly Complex Architecture for Skin Pigmentation in Africans
Abstract
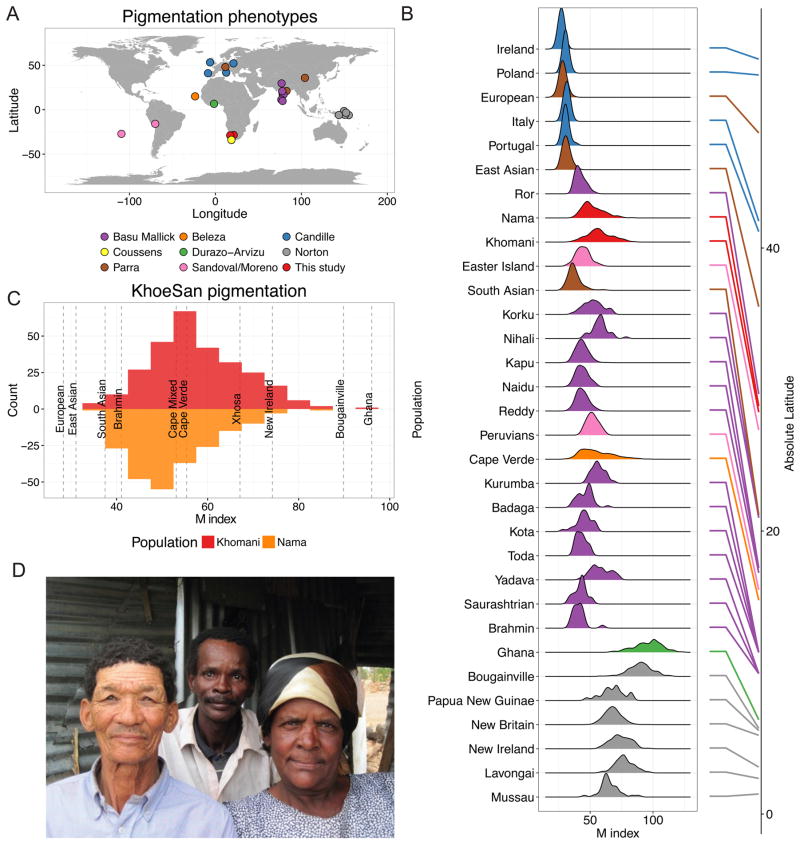
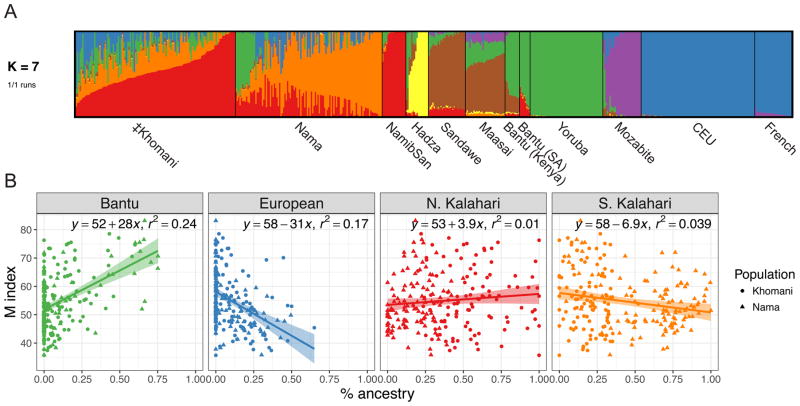
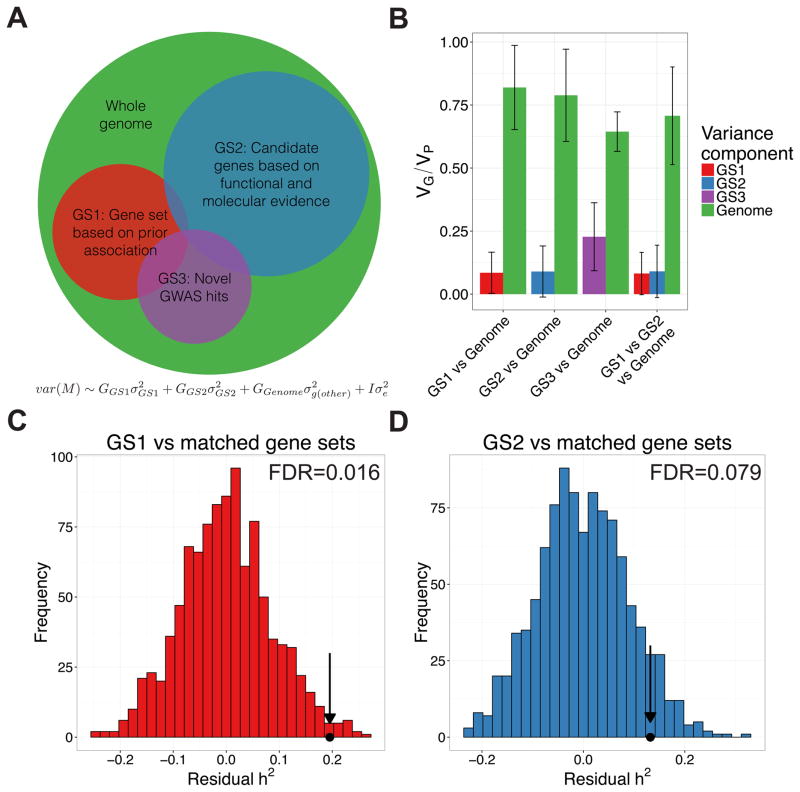
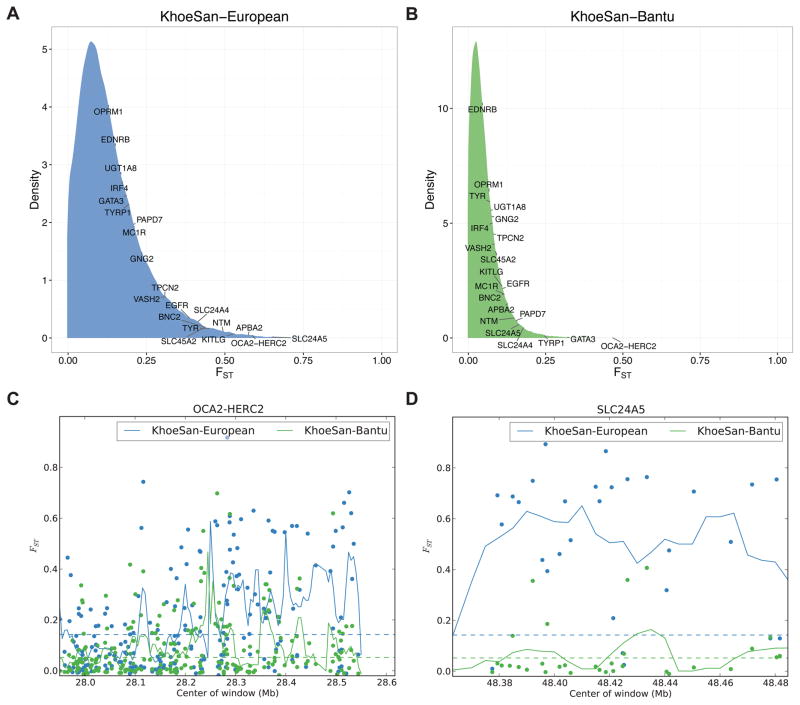
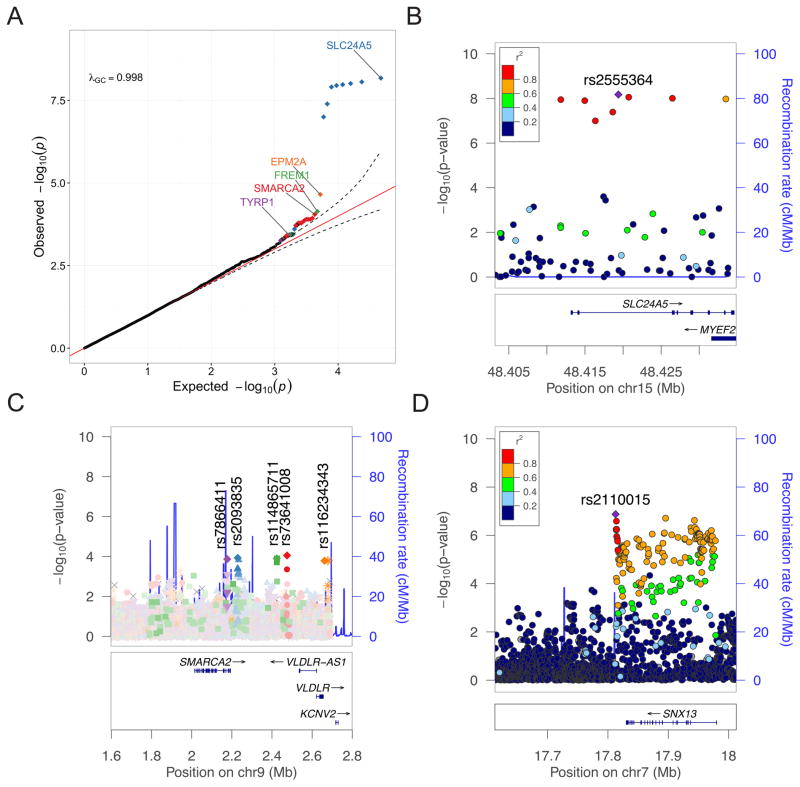
Approximately 15 genes have been directly associated with skin pigmentation variation in humans, leading to its characterization as a relatively simple trait. However, by assembling a global survey of quantitative skin pigmentation phenotypes, we demonstrate that pigmentation is more complex than previously assumed, with genetic architecture varying by latitude. We investigate polygenicity in the KhoeSan populations indigenous to southern Africa who have considerably lighter skin than equatorial Africans. We demonstrate that skin pigmentation is highly heritable, but known pigmentation loci explain only a small fraction of the variance. Rather, baseline skin pigmentation is a complex, polygenic trait in the KhoeSan. Despite this, we identify canonical and non-canonical skin pigmentation loci, including near SLC24A5, TYRP1, SMARCA2/VLDLR, and SNX13, using a genome-wide association approach complemented by targeted resequencing. By considering diverse, under-studied African populations, we show how the architecture of skin pigmentation can vary across humans subject to different local evolutionary pressures.
Keywords: Africa; heritability; human evolution; pigmentation; population genetics.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Focus on African diversity confirms complexity of skin pigmentation genetics.Genome Biol. 2018 Jan 31;19(1):13. doi: 10.1186/s13059-018-1395-3. Genome Biol. 2018. PMID: 29386055 Free PMC article.
-
Towards the full spectrum of genes for human skin colour.Pigment Cell Melanoma Res. 2018 Jul;31(4):457-458. doi: 10.1111/pcmr.12691. Epub 2018 Feb 26. Pigment Cell Melanoma Res. 2018. PMID: 29419941 No abstract available.
References
-
- Barton NH. Clines in polygenic traits. Genet Res. 1999;74:223–236. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
