

Blue light-excited LOV1 and LOV2 domains cooperatively regulate the kinase activity of full-length phototropin2 from Arabidopsis
- PMID: 29196607
- PMCID: PMC5777267
- DOI: 10.1074/jbc.RA117.000324
Blue light-excited LOV1 and LOV2 domains cooperatively regulate the kinase activity of full-length phototropin2 from Arabidopsis
Abstract
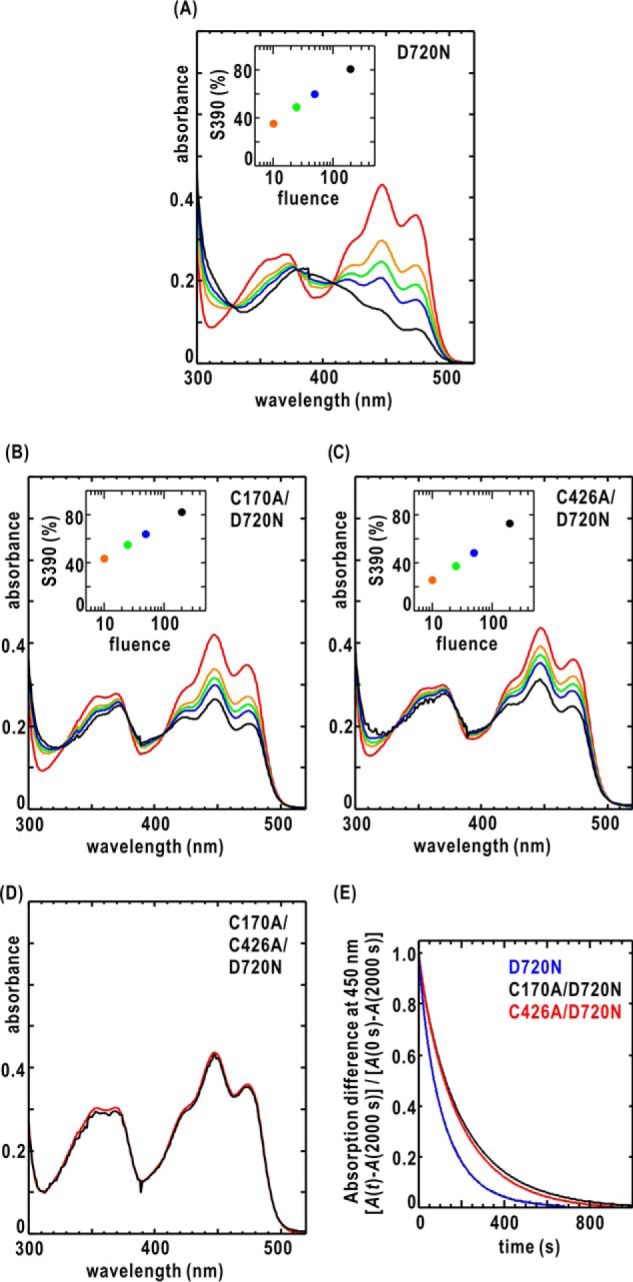
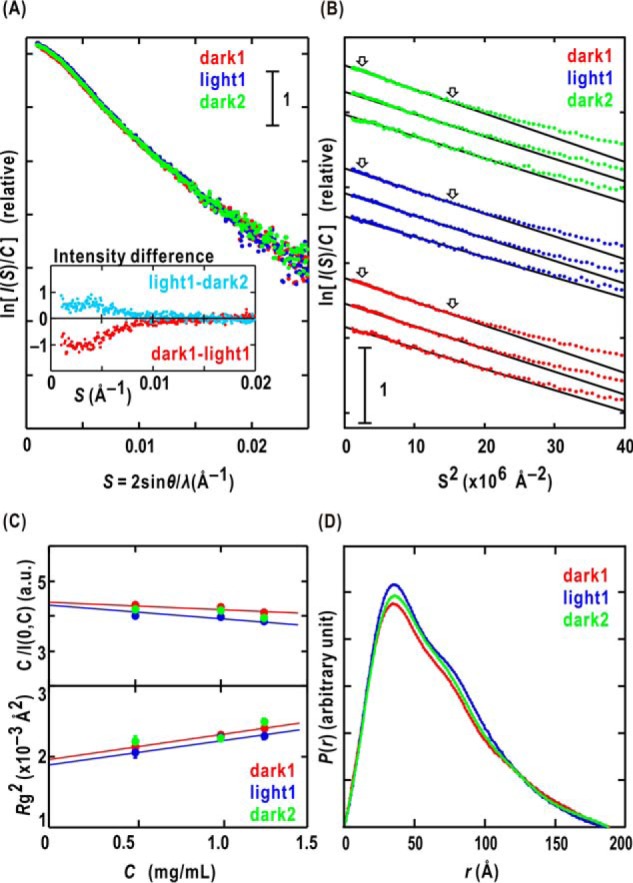
Phototropin2 (phot2) is a blue-light (BL) receptor that regulates BL-dependent activities for efficient photosynthesis in plants. phot2 comprises two BL-receiving light-oxygen-voltage-sensing domains (LOV1 and LOV2) and a kinase domain. BL-excited LOV2 is thought to be primarily responsible for the BL-dependent activation of the kinase. However, the molecular mechanisms by which small BL-induced conformational changes in the LOV2 domain are transmitted to the kinase remain unclear. Here, we used full-length wild-type and mutant phot2 proteins from Arabidopsis to study their molecular properties in the dark and under BL irradiation. Phosphorylation assays and absorption measurements indicated that the LOV1 domain assists the thermal relaxation of BL-excited LOV2 and vice versa. Using small-angle X-ray scattering and electron microscopy, we observed that phot2 forms a dimer and has a rod shape with a maximum length of 188 Å and a radius of gyration of 44 Å. Under BL, phot2 displayed large conformational changes that bent the rod shape. By superimposing the crystal structures of the LOV1 dimer, LOV2, and a homology model of the kinase to the observed changes, we inferred that the BL-dependent change consisted of positional shifts of both LOV2 and the kinase relative to LOV1. Furthermore, phot2 mutants lacking the photocycle in LOV1 or LOV2 still exhibited conformational changes under BL, suggesting that LOV1 and LOV2 cooperatively contribute to the conformational changes that activate the kinase. These results suggest that BL-activated LOV1 contributes to the kinase activity of phot2. We discuss the possible intramolecular interactions and signaling mechanisms in phot2.
Keywords: blue light; light signal transduction; photobiology; phototropin; plant biochemistry; protein kinase; small-angle X-ray scattering (SAXS); structural model.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures



References
-
- Sakai T., Kagawa T., Kasahara M., Swartz T. E., Christie J. M., Briggs W. R., Wada M., and Okada K. (2001) Arabidopsis nph1 and npl1: blue light receptors that mediate both phototropism and chloroplast relocation. Proc. Natl. Acad. Sci. U.S.A. 98, 6969–6974 10.1073/pnas.101137598 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
