

Polarity Sorting of Microtubules in the Axon
- PMID: 29198454
- PMCID: PMC5801152
- DOI: 10.1016/j.tins.2017.11.002
Polarity Sorting of Microtubules in the Axon
Abstract
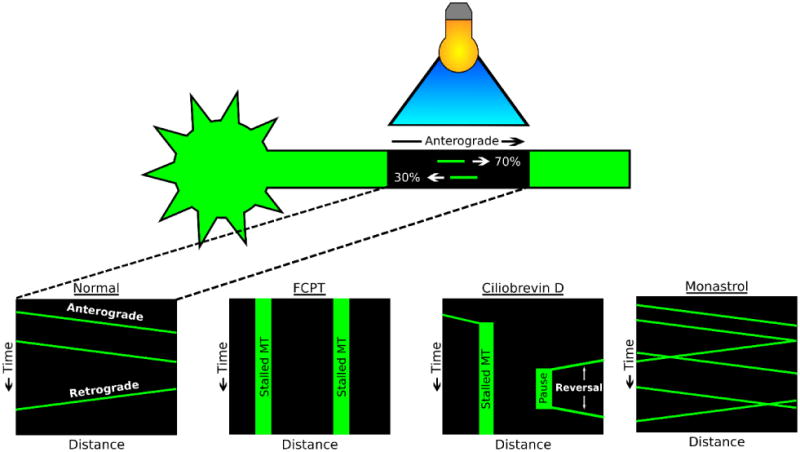
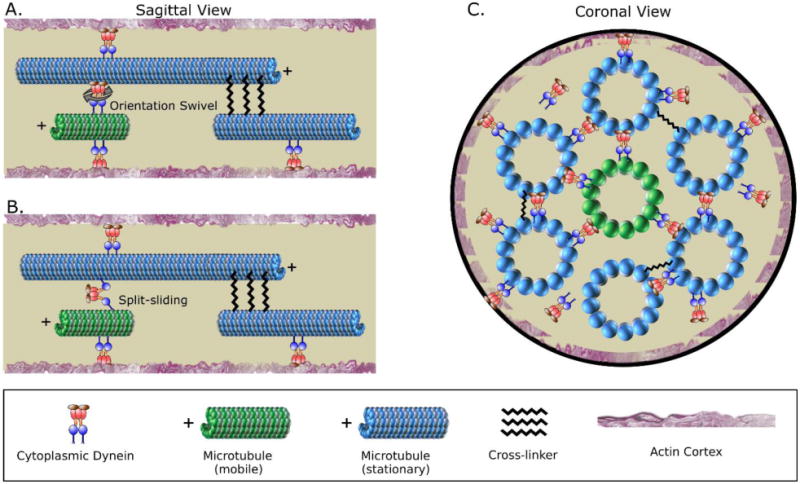
A longstanding question in cellular neuroscience is how microtubules in the axon become organized with their plus ends out, a pattern starkly different from the mixed orientation of microtubules in vertebrate dendrites. Recent attention has focused on a mechanism called polarity sorting, in which microtubules of opposite orientation are spatially separated by molecular motor proteins. Here we discuss this mechanism, and conclude that microtubules are polarity sorted in the axon by cytoplasmic dynein but that additional factors are also needed. In particular, computational modeling and experimental evidence suggest that static crosslinking proteins are required to appropriately restrict microtubule movements so that polarity sorting by cytoplasmic dynein can occur in a manner unimpeded by other motor proteins.
Keywords: axon; cytoplasmic dynein; microtubule; microtubule polarity orientation; microtubule polarity sorting; microtubule sliding.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures



References
-
- Conde C, Caceres A. Microtubule assembly, organization and dynamics in axons and dendrites. Nat Rev Neurosci. 2009;10(5):319–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
