

Anatomical Alterations in Plant Tissues Induced by Plant-Parasitic Nematodes
- PMID: 29201038
- PMCID: PMC5697168
- DOI: 10.3389/fpls.2017.01987
Anatomical Alterations in Plant Tissues Induced by Plant-Parasitic Nematodes
Abstract
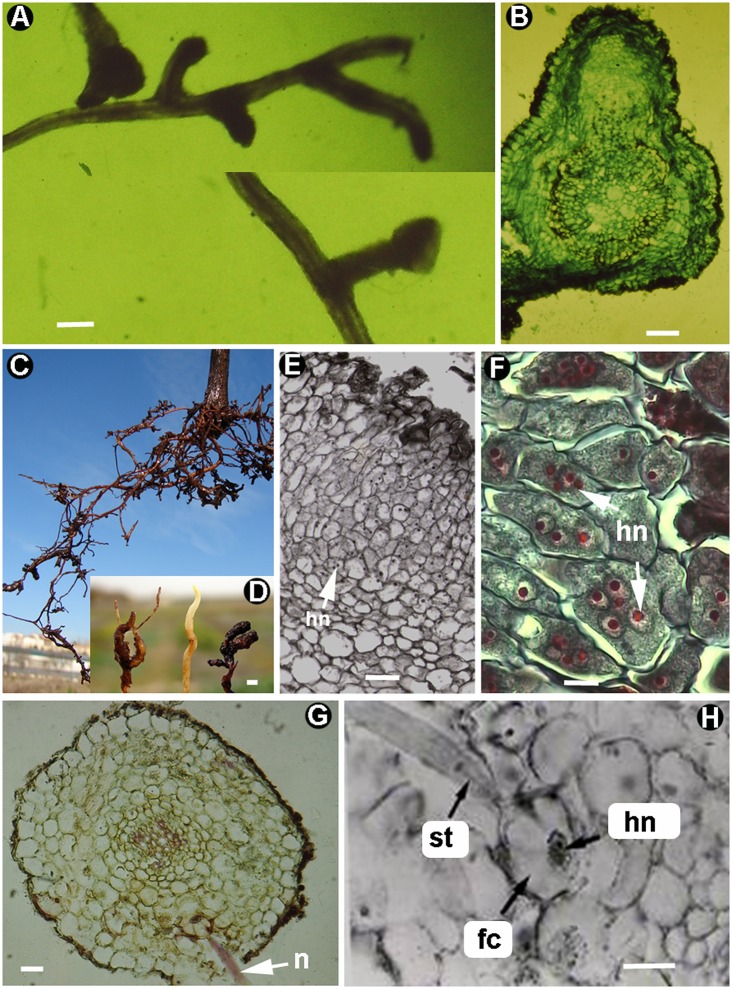
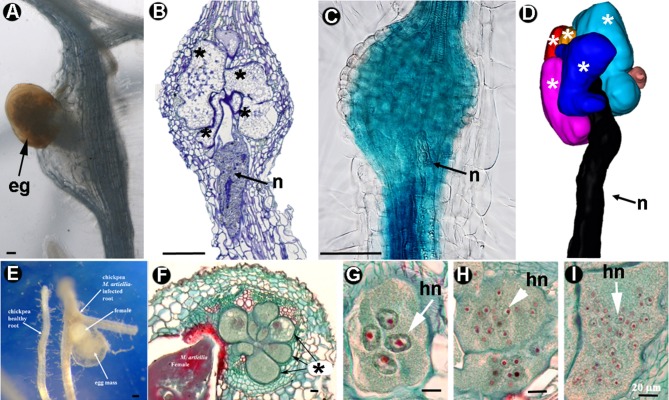
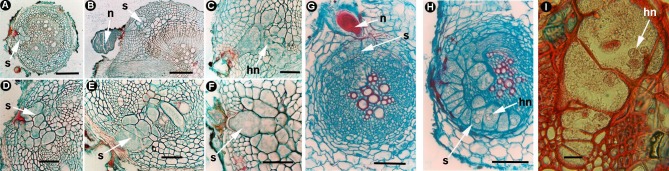
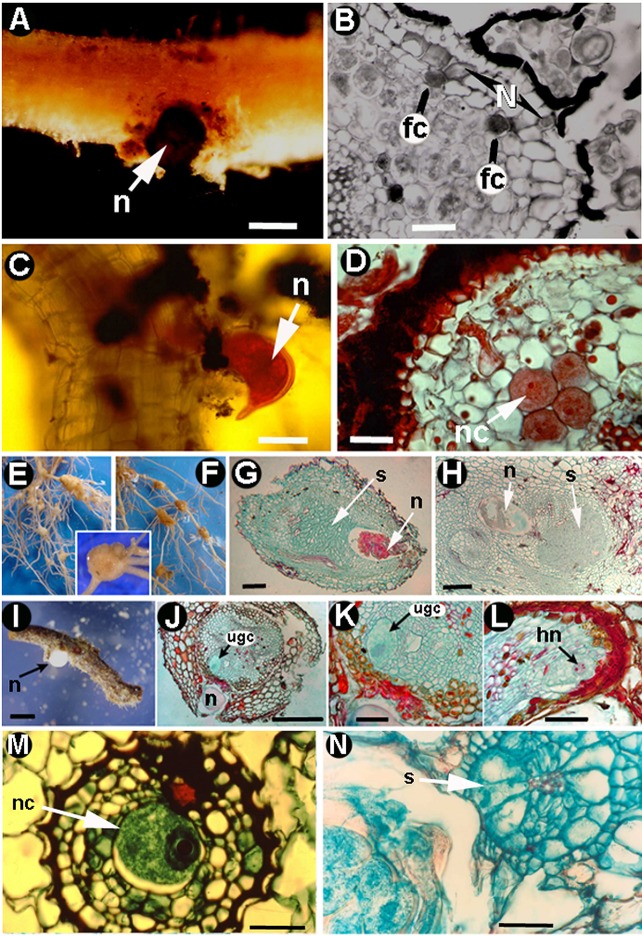
Plant-parasitic nematodes (PPNs) interact with plants in different ways, for example, through subtle feeding behavior, migrating destructively through infected tissues, or acting as virus-vectors for nepoviruses. They are all obligate biotrophic parasites as they derive their nutrients from living cells which they modify using pharyngeal gland secretions prior to food ingestion. Some of them can also shield themselves against plant defenses to sustain a relatively long lasting interaction while feeding. This paper is centered on cell types or organs that are newly induced in plants during PPN parasitism, including recent approaches to their study based on molecular biology combined with cell biology-histopathology. This issue has already been reviewed extensively for major PPNs (i.e., root-knot or cyst nematodes), but not for other genera (viz. Nacobbus aberrans, Rotylenchulus spp.). PPNs have evolved with plants and this co-evolution process has allowed the induction of new types of plant cells necessary for their parasitism. There are four basic types of feeding cells: (i) non-hypertrophied nurse cells; (ii) single giant cells; (iii) syncytia; and (iv) coenocytes. Variations in the structure of these cells within each group are also present between some genera depending on the nematode species viz. Meloidogyne or Rotylenchulus. This variability of feeding sites may be related in some way to PPN life style (migratory ectoparasites, sedentary ectoparasites, migratory ecto-endoparasites, migratory endoparasites, or sedentary endoparasites). Apart from their co-evolution with plants, the response of plant cells and roots are closely related to feeding behavior, the anatomy of the nematode (mainly stylet size, which could reach different types of cells in the plant), and the secretory fluids produced in the pharyngeal glands. These secretory fluids are injected through the stylet into perforated cells where they modify plant cytoplasm prior to food removal. Some species do not produce specialized feeding sites (viz. Ditylenchus, Subanguina), but may develop a specialized modification of the root system (e.g., unspecialized root galls or a profusion of roots). This review introduces new data on cell types and plant organs stimulated by PPNs using sources varying from traditional histopathology to new holistic methodologies.
Keywords: Ditylenchus; Globodera; Heterodera; Meloidogyne; Rotylenchulus; Xiphinema; giant cell; syncytium.
Figures

Similar articles
-
The Fight against Plant-Parasitic Nematodes: Current Status of Bacterial and Fungal Biocontrol Agents.Pathogens. 2022 Oct 13;11(10):1178. doi: 10.3390/pathogens11101178. Pathogens. 2022. PMID: 36297235 Free PMC article. Review.
-
The transcriptome of Nacobbus aberrans reveals insights into the evolution of sedentary endoparasitism in plant-parasitic nematodes.Genome Biol Evol. 2014 Aug 13;6(9):2181-94. doi: 10.1093/gbe/evu171. Genome Biol Evol. 2014. PMID: 25123114 Free PMC article.
-
Top 10 plant-parasitic nematodes in molecular plant pathology.Mol Plant Pathol. 2013 Dec;14(9):946-61. doi: 10.1111/mpp.12057. Epub 2013 Jul 1. Mol Plant Pathol. 2013. PMID: 23809086 Free PMC article. Review.
-
Co-adaptation mechanisms in plant-nematode systems.Parazitologiia. 2014 Mar-Apr;48(2):110-30. Parazitologiia. 2014. PMID: 25272462 Review.
-
Plant Proteins and Processes Targeted by Parasitic Nematode Effectors.Front Plant Sci. 2019 Jul 30;10:970. doi: 10.3389/fpls.2019.00970. eCollection 2019. Front Plant Sci. 2019. PMID: 31417587 Free PMC article. Review.
Cited by
-
Fungal and Bacterial Endophytes as Microbial Control Agents for Plant-Parasitic Nematodes.Int J Environ Res Public Health. 2021 Apr 17;18(8):4269. doi: 10.3390/ijerph18084269. Int J Environ Res Public Health. 2021. PMID: 33920580 Free PMC article. Review.
-
In vitro evaluation of Penicillium chrysogenum Snef1216 against Meloidogyne incognita (root-knot nematode).Sci Rep. 2020 May 20;10(1):8342. doi: 10.1038/s41598-020-65262-z. Sci Rep. 2020. PMID: 32433576 Free PMC article.
-
A review on fungal endophytes of the family Fabaceae, their metabolic diversity and biological applications.Heliyon. 2025 Jan 23;11(3):e42153. doi: 10.1016/j.heliyon.2025.e42153. eCollection 2025 Feb 15. Heliyon. 2025. PMID: 40196783 Free PMC article. Review.
-
In silico secretome analyses of the polyphagous root-knot nematode Meloidogyne javanica: a resource for studying M. javanica secreted proteins.BMC Genomics. 2023 Jun 1;24(1):296. doi: 10.1186/s12864-023-09366-6. BMC Genomics. 2023. PMID: 37264326 Free PMC article.
-
A Deep Learning-Based Decision Support Tool for Plant-Parasitic Nematode Management.J Imaging. 2023 Nov 6;9(11):240. doi: 10.3390/jimaging9110240. J Imaging. 2023. PMID: 37998089 Free PMC article.
References
-
- Agrios G. N. (2005). Plant Pathology. London: Elsevier, Academic Press.
-
- Ali N., Tavoillot J., Mateille T., Chapuis E., Besnard G., El Bakkali A., et al. (2015). A new root-knot nematode Meloidogyne spartelensis n. sp. (Nematoda: Meloidogynidae) in Northern Morocco. Eur. J. Plant Pathol. 143, 25–42. 10.1007/s10658-015-0662-3 - DOI
-
- Back M. A., Haydock P. P. J., Jenkinson P. (2002). Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathol. 51, 683–697. 10.1046/j.1365-3059.2002.00785.x - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous