

Transmissible Gastroenteritis Virus Papain-Like Protease 1 Antagonizes Production of Interferon- β through Its Deubiquitinase Activity
- PMID: 29201911
- PMCID: PMC5672592
- DOI: 10.1155/2017/7089091
Transmissible Gastroenteritis Virus Papain-Like Protease 1 Antagonizes Production of Interferon- β through Its Deubiquitinase Activity
Abstract
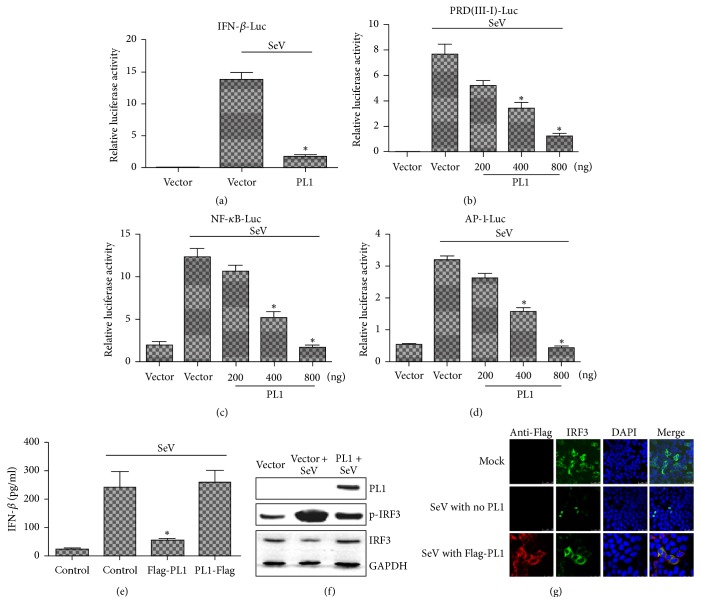
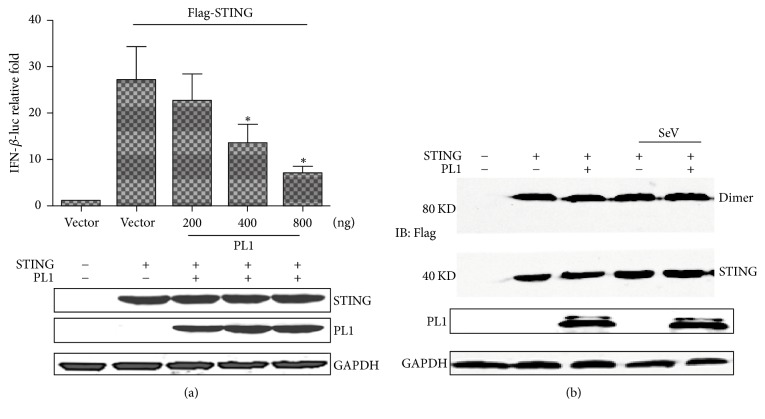
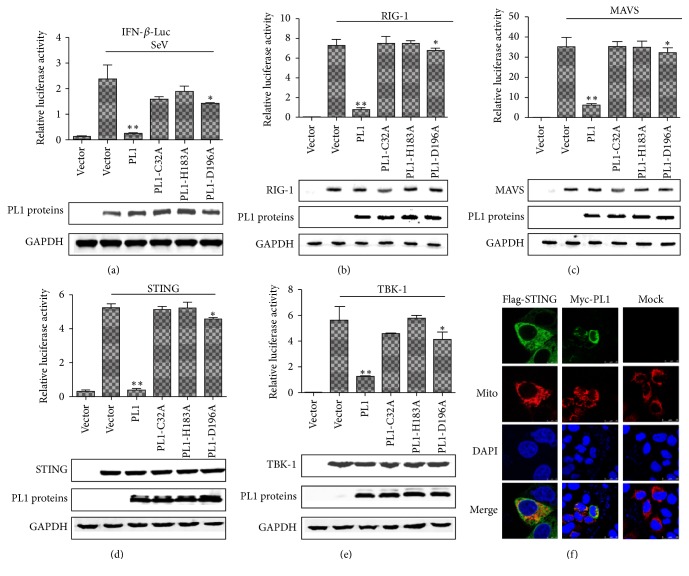
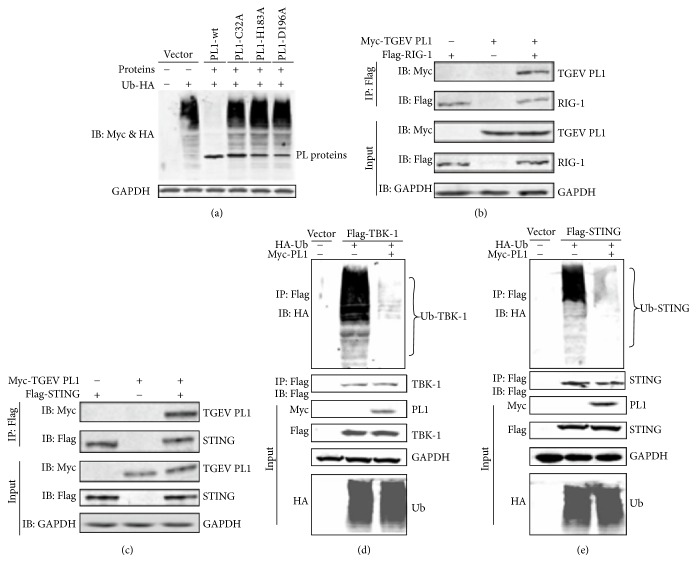
Coronaviruses (CoVs), such as human coronavirus NL63 (HCoV-NL63), severe acute respiratory syndrome CoV (SARS-CoV), murine hepatitis virus (MHV), porcine epidemic diarrhea virus (PEDV), and Middle East Respiratory Syndrome Coronavirus (MERS-CoV), encode papain-like (PL) proteases that inhibit Sendai virus- (SeV-) induced interferon (IFN-β) production. Recently, the crystal structure of transmissible gastroenteritis virus (TGEV) PL1 has been solved, which was similar to that of SARS-CoV PL2pro, which may antagonize host innate immunity. However, very little is known about whether TGEV PL1 can antagonize host innate immune response. Here, we presented evidence that TGEV PL1 encoded by the replicase gene could suppress the IFN-β expression and inhibit the nuclear translocation of interferon regulatory factor 3 (IRF3). The ability to antagonize IFN-β production was dependent on the intact catalytic activity of PL1. Furthermore, TGEV PL1 exerted deubiquitinase (DUB) activity which strongly inhibited the retinoic acid-induced gene I- (RIG-1-) and stimulator of interferon gene- (STING-) dependent IFN expression. Our data collectively suggest that TGEV PL1 can inhibit the IFN-β expression and interfere with RIG-1- and STING-mediated signaling through a viral DUB activity. Our study has yielded strong evidence for the TGEV PL1 mechanisms that counteract the host innate immunity.
Figures
Similar articles
-
Papain-like protease 1 from transmissible gastroenteritis virus: crystal structure and enzymatic activity toward viral and cellular substrates.J Virol. 2010 Oct;84(19):10063-73. doi: 10.1128/JVI.00898-10. Epub 2010 Jul 28. J Virol. 2010. PMID: 20668092 Free PMC article.
-
The papain-like protease of porcine epidemic diarrhea virus negatively regulates type I interferon pathway by acting as a viral deubiquitinase.J Gen Virol. 2013 Jul;94(Pt 7):1554-1567. doi: 10.1099/vir.0.051169-0. Epub 2013 Apr 17. J Gen Virol. 2013. PMID: 23596270 Free PMC article.
-
Proteolytic processing, deubiquitinase and interferon antagonist activities of Middle East respiratory syndrome coronavirus papain-like protease.J Gen Virol. 2014 Mar;95(Pt 3):614-626. doi: 10.1099/vir.0.059014-0. Epub 2013 Dec 20. J Gen Virol. 2014. PMID: 24362959
-
Immune evasion of porcine enteric coronaviruses and viral modulation of antiviral innate signaling.Virus Res. 2016 Dec 2;226:128-141. doi: 10.1016/j.virusres.2016.05.015. Epub 2016 May 19. Virus Res. 2016. PMID: 27212682 Free PMC article. Review.
-
Modulation of Innate Antiviral Immune Response by Porcine Enteric Coronavirus.Front Microbiol. 2022 Feb 14;13:845137. doi: 10.3389/fmicb.2022.845137. eCollection 2022. Front Microbiol. 2022. PMID: 35237253 Free PMC article. Review.
Cited by
-
Interplay between RNA viruses and cGAS/STING axis in innate immunity.Front Cell Infect Microbiol. 2023 Apr 3;13:1172739. doi: 10.3389/fcimb.2023.1172739. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37077526 Free PMC article. Review.
-
Interactions between SARS coronavirus 2 papain-like protease and immune system: A potential drug target for the treatment of COVID-19.Scand J Immunol. 2021 Oct;94(4):e13044. doi: 10.1111/sji.13044. Epub 2021 Aug 10. Scand J Immunol. 2021. PMID: 33872387 Free PMC article. Review.
-
Differential induction of type I interferon by type I and type II feline coronaviruses in vitro.Res Vet Sci. 2018 Oct;120:57-62. doi: 10.1016/j.rvsc.2018.09.002. Epub 2018 Sep 21. Res Vet Sci. 2018. PMID: 30265871 Free PMC article.
-
Porcine deltacoronavirus nonstructural protein 2 inhibits type I and III IFN production by targeting STING for degradation.Vet Res. 2024 Jun 17;55(1):79. doi: 10.1186/s13567-024-01330-w. Vet Res. 2024. PMID: 38886840 Free PMC article.
-
Suppression of NF-κB Activity: A Viral Immune Evasion Mechanism.Viruses. 2018 Aug 4;10(8):409. doi: 10.3390/v10080409. Viruses. 2018. PMID: 30081579 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
