

PnuT uses a facilitated diffusion mechanism for thiamine uptake
- PMID: 29203477
- PMCID: PMC5749112
- DOI: 10.1085/jgp.201711850
PnuT uses a facilitated diffusion mechanism for thiamine uptake
Abstract
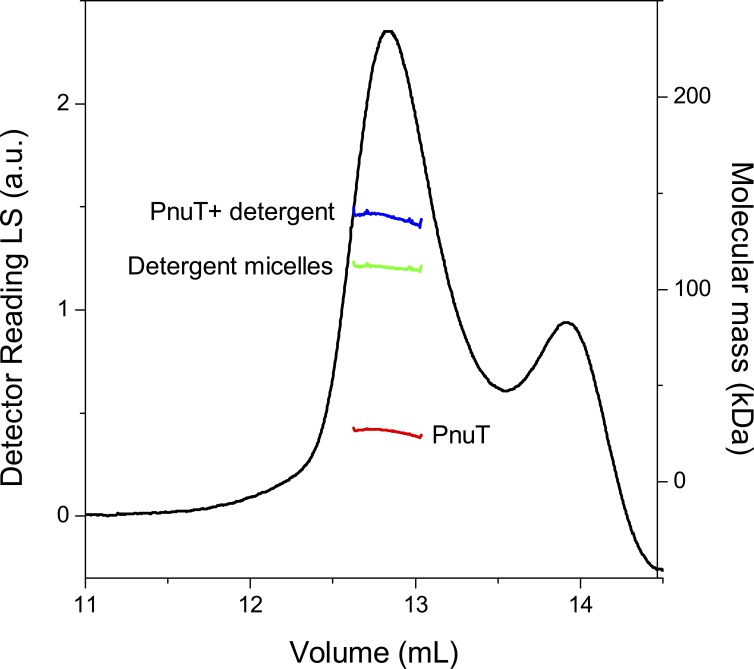
Membrane transporters of the bacterial pyridine nucleotide uptake (Pnu) family mediate the uptake of various B-type vitamins. For example, the PnuT transporters have specificity for vitamin B1 (thiamine). It has been hypothesized that Pnu transporters are facilitators that allow passive transport of the vitamin substrate across the membrane. Metabolic trapping by phosphorylation would then lead to accumulation of the transported substrates in the cytoplasm. However, experimental evidence for such a transport mechanism is lacking. Here, to determine the mechanism of thiamine transport, we purify PnuTSw from Shewanella woodyi and reconstitute it in liposomes to determine substrate binding and transport properties. We show that the electrochemical gradient of thiamine solely determines the direction of transport, consistent with a facilitated diffusion mechanism. Further, PnuTSw can bind and transport thiamine as well as the thiamine analogues pyrithiamine and oxythiamine, but does not recognize the phosphorylated derivatives thiamine monophosphate and thiamine pyrophosphate as substrates, consistent with a metabolic trapping mechanism. Guided by the crystal structure of the homologous nicotinamide riboside transporter PnuC, we perform mutagenesis experiments, which reveal residues involved in substrate binding and gating. The facilitated diffusion mechanism of transport used by PnuTSw contrasts sharply with the active transport mechanisms used by other bacterial thiamine transporters.
© 2018 Jaehme et al.
Figures






Similar articles
-
Thiamin transport in Helicobacter pylori lacking the de novo synthesis of thiamin.Microbiology (Reading). 2019 Feb;165(2):224-232. doi: 10.1099/mic.0.000765. Epub 2019 Jan 8. Microbiology (Reading). 2019. PMID: 30620266
-
Structure, function, evolution, and application of bacterial Pnu-type vitamin transporters.Biol Chem. 2015 Sep;396(9-10):955-66. doi: 10.1515/hsz-2015-0113. Biol Chem. 2015. PMID: 26352203 Review.
-
Directed Evolution of Membrane Transport Using Synthetic Selections.ACS Synth Biol. 2018 Mar 16;7(3):789-793. doi: 10.1021/acssynbio.7b00407. Epub 2018 Feb 28. ACS Synth Biol. 2018. PMID: 29474058
-
Both thiamine uptake and biosynthesis of thiamine precursors are required for intracellular replication of Listeria monocytogenes.J Bacteriol. 2009 Apr;191(7):2218-27. doi: 10.1128/JB.01636-08. Epub 2009 Jan 30. J Bacteriol. 2009. PMID: 19181806 Free PMC article.
-
The twisted relation between Pnu and SWEET transporters.Trends Biochem Sci. 2015 Apr;40(4):183-8. doi: 10.1016/j.tibs.2015.02.002. Epub 2015 Mar 7. Trends Biochem Sci. 2015. PMID: 25757400 Review.
Cited by
-
Structural and Functional Characterization of NadR from Lactococcus lactis.Molecules. 2020 Apr 22;25(8):1940. doi: 10.3390/molecules25081940. Molecules. 2020. PMID: 32331317 Free PMC article.
-
Mitochondrial transport and metabolism of the vitamin B-derived cofactors thiamine pyrophosphate, coenzyme A, FAD and NAD+ , and related diseases: A review.IUBMB Life. 2022 Jul;74(7):592-617. doi: 10.1002/iub.2612. Epub 2022 Mar 18. IUBMB Life. 2022. PMID: 35304818 Free PMC article. Review.
-
Vitamin biosynthesis in the gut: interplay between mammalian host and its resident microbiota.Microbiol Mol Biol Rev. 2025 Jun 25;89(2):e0018423. doi: 10.1128/mmbr.00184-23. Epub 2025 Apr 2. Microbiol Mol Biol Rev. 2025. PMID: 40172109 Review.
-
A second riboswitch class for the enzyme cofactor NAD.RNA. 2021 Jan;27(1):99-105. doi: 10.1261/rna.077891.120. Epub 2020 Oct 21. RNA. 2021. PMID: 33087526 Free PMC article.
-
Carriers, exchangers, and cotransporters in the first 100 years of the Journal of General Physiology.J Gen Physiol. 2018 Aug 6;150(8):1063-1080. doi: 10.1085/jgp.201812078. Epub 2018 Jul 20. J Gen Physiol. 2018. PMID: 30030301 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
