

Crystal and solution structures of human protein-disulfide isomerase-like protein of the testis (PDILT) provide insight into its chaperone activity
- PMID: 29203529
- PMCID: PMC5787798
- DOI: 10.1074/jbc.M117.797290
Crystal and solution structures of human protein-disulfide isomerase-like protein of the testis (PDILT) provide insight into its chaperone activity
Abstract
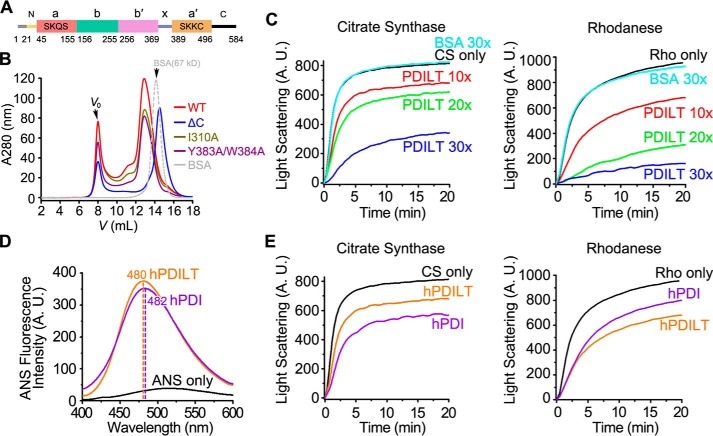
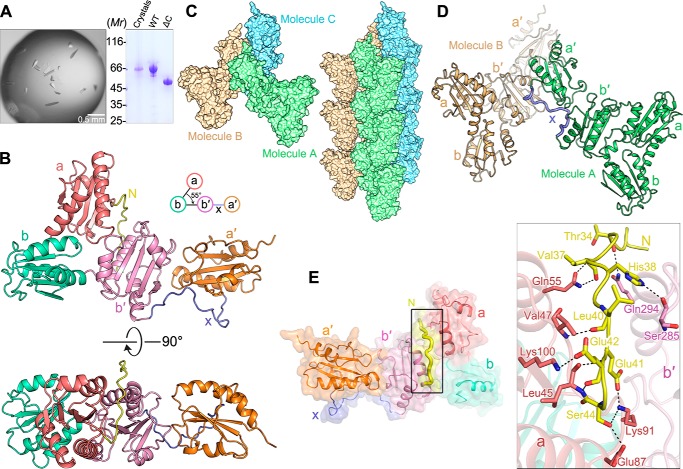
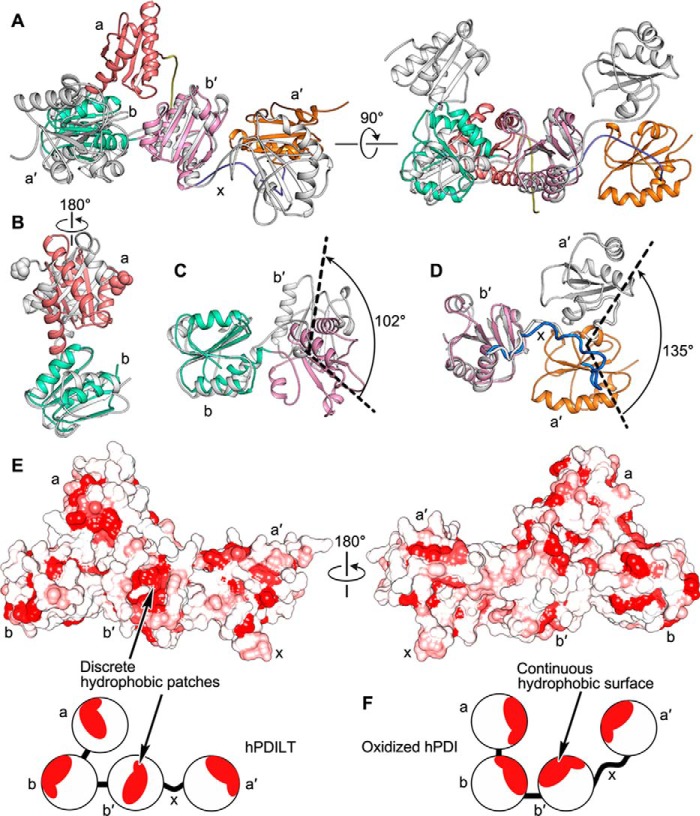
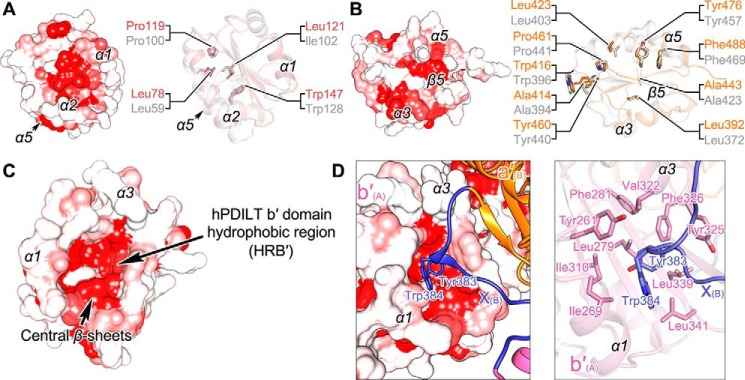
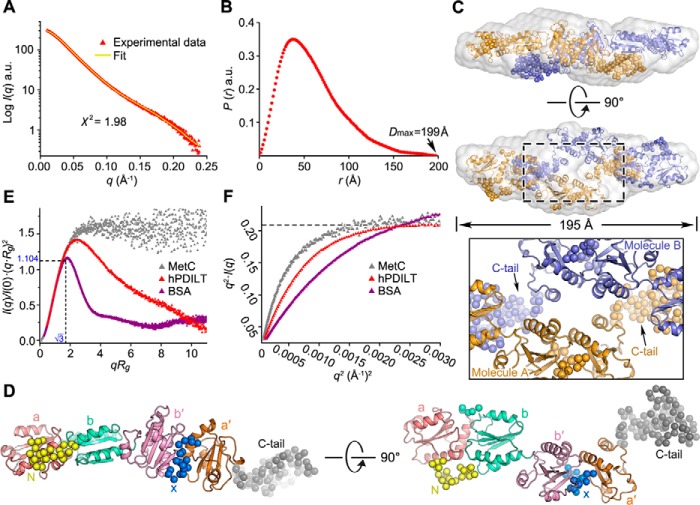
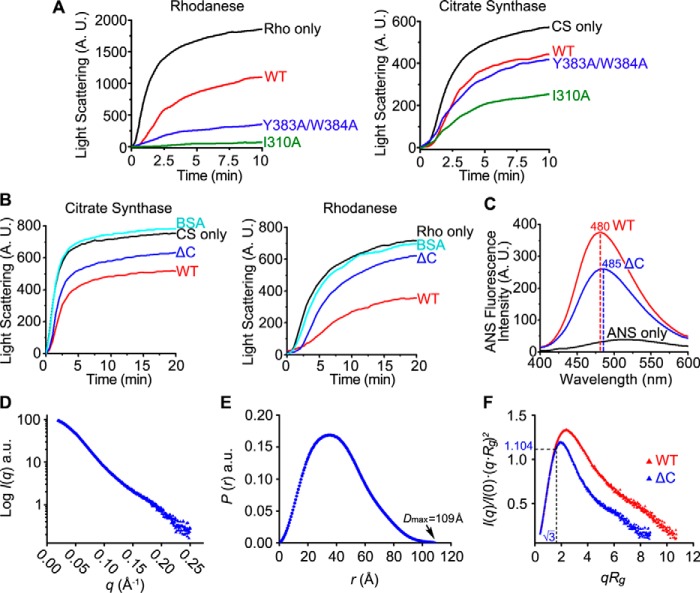
Protein-disulfide isomerase-like protein of the testis (PDILT), a member of the protein-disulfide isomerase family, is a chaperone essential for the folding of spermatogenesis-specific proteins in male postmeiotic germ cells. However, the structural mechanisms that regulate the chaperone function of PDILTs are unknown. Here, we report the structures of human PDILT (hPDILT) determined by X-ray crystallography to 2.4 Å resolution and small-angle X-ray scattering (SAXS). Distinct from previously reported U-like structures of related PDI family proteins, our structures revealed that hPDILT folds into a compact L-like structure in crystals and into an extended chain-like structure in solution. The hydrophobic regions and the hydrophobic pockets in hPDILT, which are important for substrate recognition, were clearly delineated in the crystal structure. Moreover, our results of the SAXS analysis and of structure-based substitutions and truncations indicated that the C-terminal tail in hPDILT is required for suppression of aggregation of denatured proteins, suggesting that the tail is crucial for the chaperone activity of PDILT. Taken together, our findings have identified the critical regions and conformational changes of PDILT that enable and control its activity. These results advance our understanding of the structural mechanisms involved in the chaperone activity of PDILT.
Keywords: X-ray crystallography; conformational change; molecular chaperone; protein structure; small-angle X-ray scattering (SAXS).
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
