

Review
doi: 10.1104/pp.17.01522.
Epub 2017 Dec 4.
Membrane Dynamics and Multiple Functions of Oil Bodies in Seeds and Leaves
Affiliations
- PMID: 29203559
- PMCID: PMC5761825
- DOI: 10.1104/pp.17.01522
Item in Clipboard
Review
Membrane Dynamics and Multiple Functions of Oil Bodies in Seeds and Leaves
Plant Physiol.
2018 Jan.
Abstract
Oil bodies have multiple functions: oleosin-mediated freezing tolerance of seeds, direct interaction with glyoxysomes for lipid degradation in seedlings, and antifungal compound production in leaves.
Figures
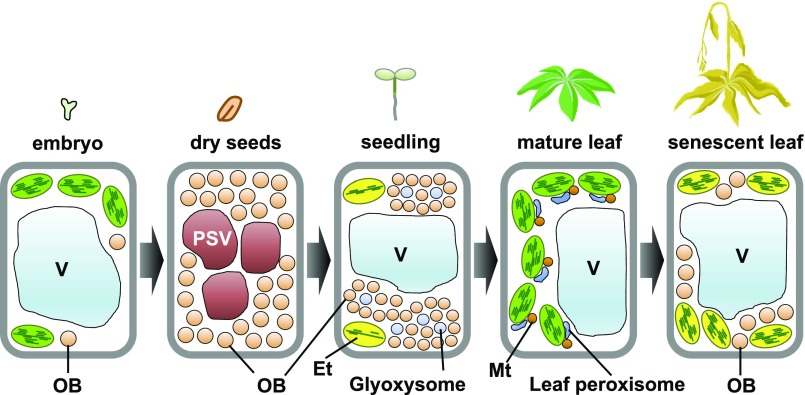
Schematic view of oil body (OB) formation and degradation during plant development. Oil bodies form in embryos and are abundant in dry seeds. During seed germination and subsequent seedling growth, glyoxysomal enzymes degrade oil bodies to release stored energy. Mature leaves contain few oil bodies, whereas oil bodies accumulate in senescent leaves. Et, Etioplast; Mt, mitochondria; PSV, protein storage vacuole; V, vacuole.
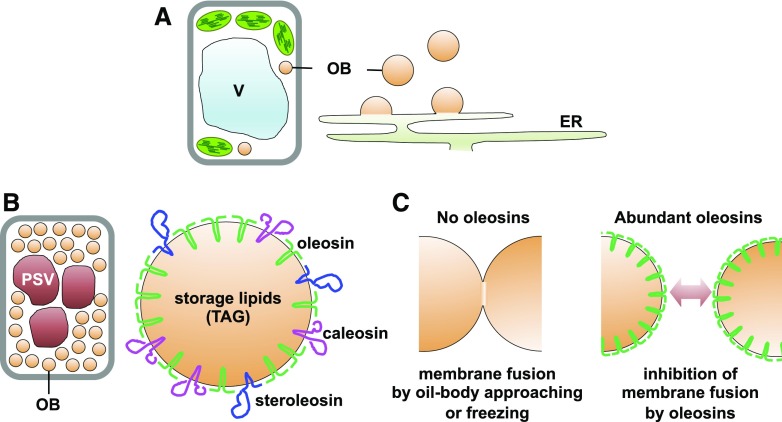
Specific proteins accumulate on the oil body (OB) membrane surface, and oleosins prevent oil bodies from fusing with each other in seeds. A, Oil bodies bud off from the ER. B, Seed oil bodies contain three membrane proteins: oleosin, caleosin, and steroleosin. Oleosins are the most abundant oil body membrane protein. C, Oleosins function to inhibit oil body fusion. Oleosin deficiency causes oil body fusion (left), whereas high expression of oleosin prevents oil body fusion (right). TAG, Triacylglycerol; V, vacuole.
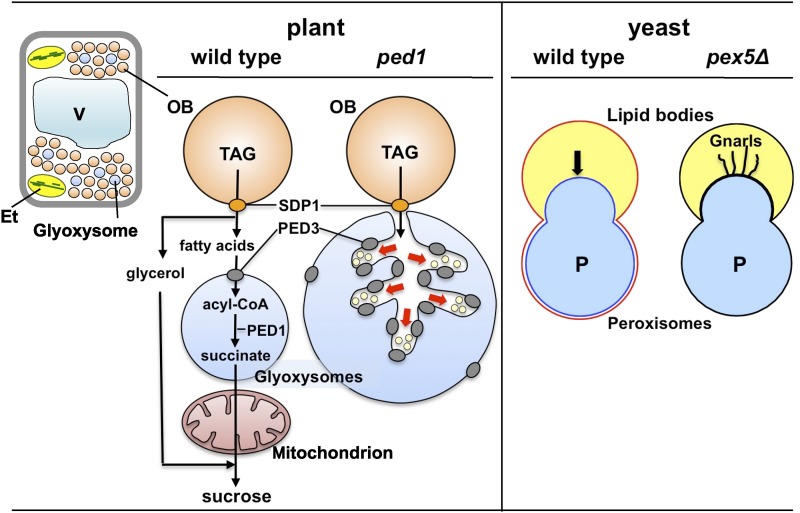
The ped1 mutant reveals communication between oil bodies (OB) and glyoxysomes. Triacylglycerol (TAG) stored in oil bodies is catabolized during seedling heterotrophic growth. TAG is hydrolyzed by triacylglycerol lipase (SDP1) to produce glycerol and free fatty acids, which are ultimately converted into Suc. Suc is metabolized further to support seedling growth. Free fatty acids are imported into glyoxysomes by PED3, which is localized on the glyoxysomal membrane, and are subsequently activated to acyl-CoA forms. Then, acyl-CoA is converted to succinate in the glyoxysome via fatty acid β-oxidation and the glyoxylate cycle. PED1 is a 3-ketoacyl-CoA thiolase that catalyzes the last step of fatty acid β-oxidation. Succinate is converted into Suc by successive enzymatic reactions in mitochondria and the cytosol. Aberrantly enlarged glyoxysome morphologies are observed in ped1 seedlings. Dendritic tubules are formed by glyoxysomal membrane invaginations at the sites of oil body-glyoxysome contact. These tubules encompass small vesicles that may be derived from the oil body (red arrows). PED3 is significantly more enriched on the dendritic tubule membrane than on the outer glyoxysomal membrane. In yeast, peroxisomes invade and stably fuse with the lipid body. The lipid body monolayer membrane fuses with the peroxisomal membrane outer leaflet to form a single outer leaflet (red line), whereas the peroxisomal membrane inner leaflet still surrounds the peroxisomal matrix (blue line). Pexopodia invade the lipid body (arrow). In the yeast pex5Δ mutant, the lipid body often forms elongated, curled, and tangled tubules called gnarls. Et, Etioplast; V, vacuole.
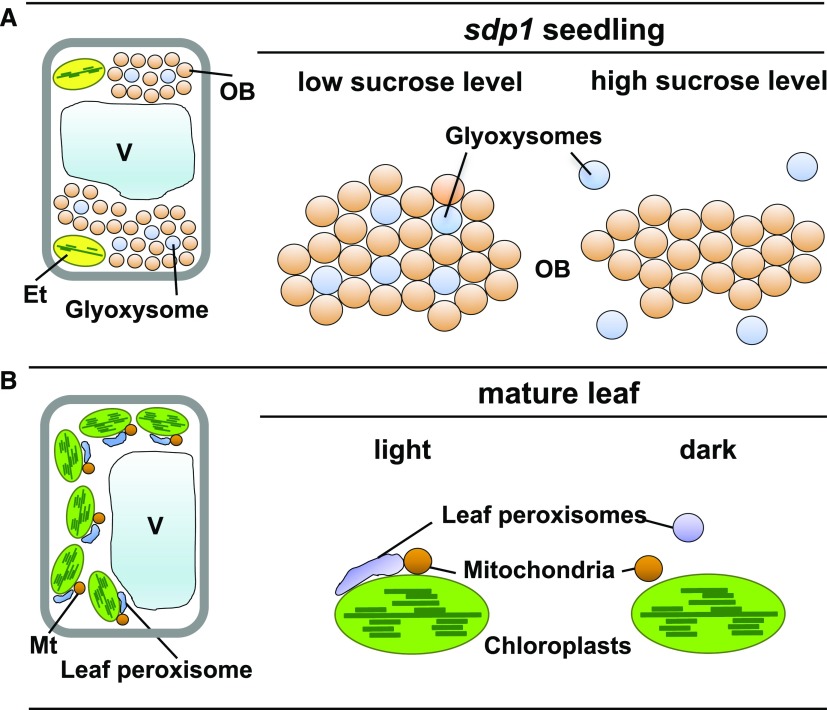
Glyoxysomes and leaf peroxisomes differentially associate and disassociate with other organelles under specific conditions. A, Oil body (OB)-glyoxysome interactions in sdp1 mutant seedlings at the heterotrophic growth stage. When sdp1 seedlings are grown without Suc supplementation, many oil bodies and glyoxysomes associate physically with each other to form a mixed aggregation. By contrast, glyoxysomes dissociate from the aggregates when sdp1 seedlings are grown in the presence of exogenously supplied Suc. B, Interactions between leaf peroxisomes, chloroplasts, and mitochondria (Mt) in photosynthetic cells of mature leaves. Under light conditions (light), elongated leaf peroxisomes associate with chloroplasts and mitochondria. Under dark conditions (dark), leaf peroxisomes become spherical and dissociate from chloroplasts. Et, Etioplast; V, vacuole.
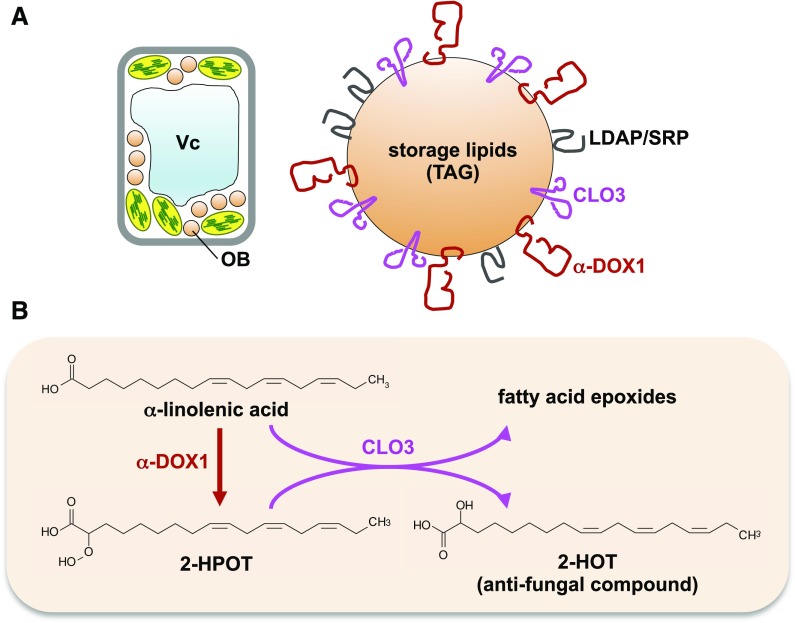
Leaf oil body (OB) proteins and their functions. A, Oil bodies in senescent leaves contain CLO3, α-DOX1, and LDAP/SRPP proteins. B, CLO3 and α-DOX1 cooperatively biosynthesize the antifungal compound 2-HOT on the oil body membrane. 2-HPOT, 2-Hydroperoxy-octadecatrienoic acid; TAG, triacylglycerol; Vc, vacuole.


Similar articles
-
Leaf oil bodies are subcellular factories producing antifungal oxylipins.Curr Opin Plant Biol. 2015 Jun;25:145-50. doi: 10.1016/j.pbi.2015.05.019. Epub 2015 Jun 5. Curr Opin Plant Biol. 2015. PMID: 26051035 Review.
-
Sterols are required for the coordinated assembly of lipid droplets in developing seeds.Nat Commun. 2021 Sep 22;12(1):5598. doi: 10.1038/s41467-021-25908-6. Nat Commun. 2021. PMID: 34552075 Free PMC article.
-
Lipid Droplets: Packing Hydrophobic Molecules Within the Aqueous Cytoplasm.Annu Rev Plant Biol. 2023 May 22;74:195-223. doi: 10.1146/annurev-arplant-070122-021752. Epub 2022 Nov 22. Annu Rev Plant Biol. 2023. PMID: 36413579 Review.
-
Arabidopsis lipid droplet-associated protein (LDAP) - interacting protein (LDIP) influences lipid droplet size and neutral lipid homeostasis in both leaves and seeds.Plant J. 2017 Dec;92(6):1182-1201. doi: 10.1111/tpj.13754. Epub 2017 Nov 27. Plant J. 2017. PMID: 29083105
-
Arabidopsis SEIPIN Proteins Modulate Triacylglycerol Accumulation and Influence Lipid Droplet Proliferation.Plant Cell. 2015 Sep;27(9):2616-36. doi: 10.1105/tpc.15.00588. Epub 2015 Sep 11. Plant Cell. 2015. PMID: 26362606 Free PMC article.
Cited by
-
Lipid Droplet Isolation from Arabidopsis thaliana Leaves.Bio Protoc. 2020 Dec 20;10(24):e3867. doi: 10.21769/BioProtoc.3867. eCollection 2020 Dec 20. Bio Protoc. 2020. PMID: 33659507 Free PMC article.
-
Bacillus subtilis biofilm matrix components target seed oil bodies to promote growth and anti-fungal resistance in melon.Nat Microbiol. 2022 Jul;7(7):1001-1015. doi: 10.1038/s41564-022-01134-8. Epub 2022 Jun 6. Nat Microbiol. 2022. PMID: 35668112 Free PMC article.
-
Seeds as oil factories.Plant Reprod. 2018 Sep;31(3):213-235. doi: 10.1007/s00497-018-0325-6. Epub 2018 Feb 10. Plant Reprod. 2018. PMID: 29429143 Review.
-
Polyamine depletion enhances oil body mobilization through possible regulation of oleosin degradation and aquaporin abundance on its membrane.Plant Signal Behav. 2023 Dec 31;18(1):2217027. doi: 10.1080/15592324.2023.2217027. Plant Signal Behav. 2023. PMID: 37243675 Free PMC article.
-
Plant Unsaturated Fatty Acids: Multiple Roles in Stress Response.Front Plant Sci. 2020 Sep 4;11:562785. doi: 10.3389/fpls.2020.562785. eCollection 2020. Front Plant Sci. 2020. PMID: 33013981 Free PMC article. Review.
References
-
- Abell BM, Hahn M, Holbrook LA, Moloney MM (2004) Membrane topology and sequence requirements for oil body targeting of oleosin. Plant J 37: 461–470 - PubMed
-
- Abell BM, High S, Moloney MM (2002) Membrane protein topology of oleosin is constrained by its long hydrophobic domain. J Biol Chem 277: 8602–8610 - PubMed
-
- Avila CA, Arevalo-Soliz LM, Lorence A, Goggin FL (2013) Expression of α-DIOXYGENASE 1 in tomato and Arabidopsis contributes to plant defenses against aphids. Mol Plant Microbe Interact 26: 977–986 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
