

A bifunctional cellulase-xylanase of a new Chryseobacterium strain isolated from the dung of a straw-fed cattle
- PMID: 29205864
- PMCID: PMC5812240
- DOI: 10.1111/1751-7915.13034
A bifunctional cellulase-xylanase of a new Chryseobacterium strain isolated from the dung of a straw-fed cattle
Abstract
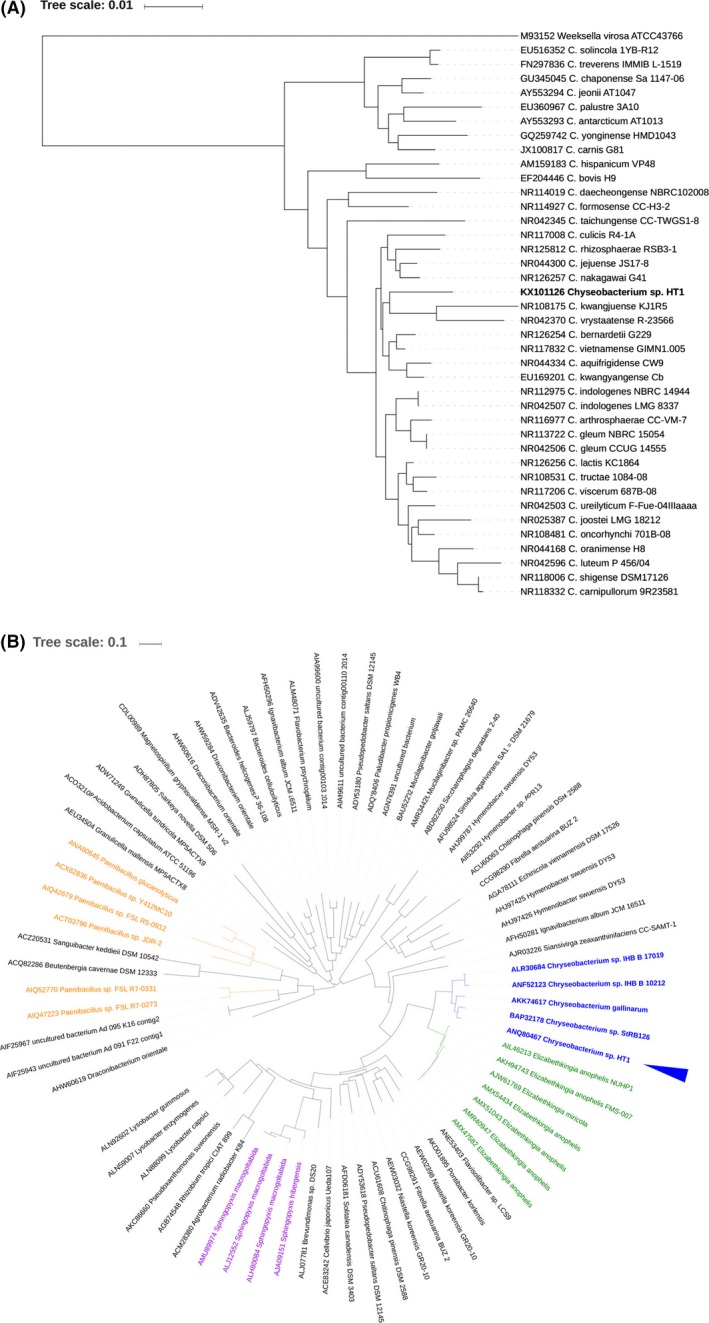
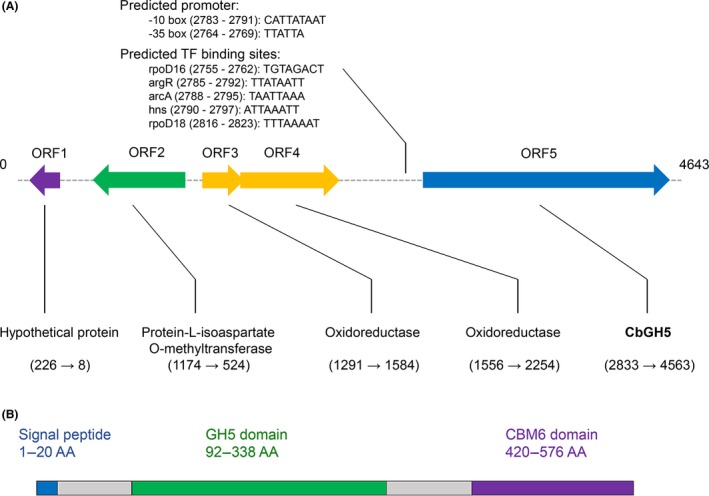
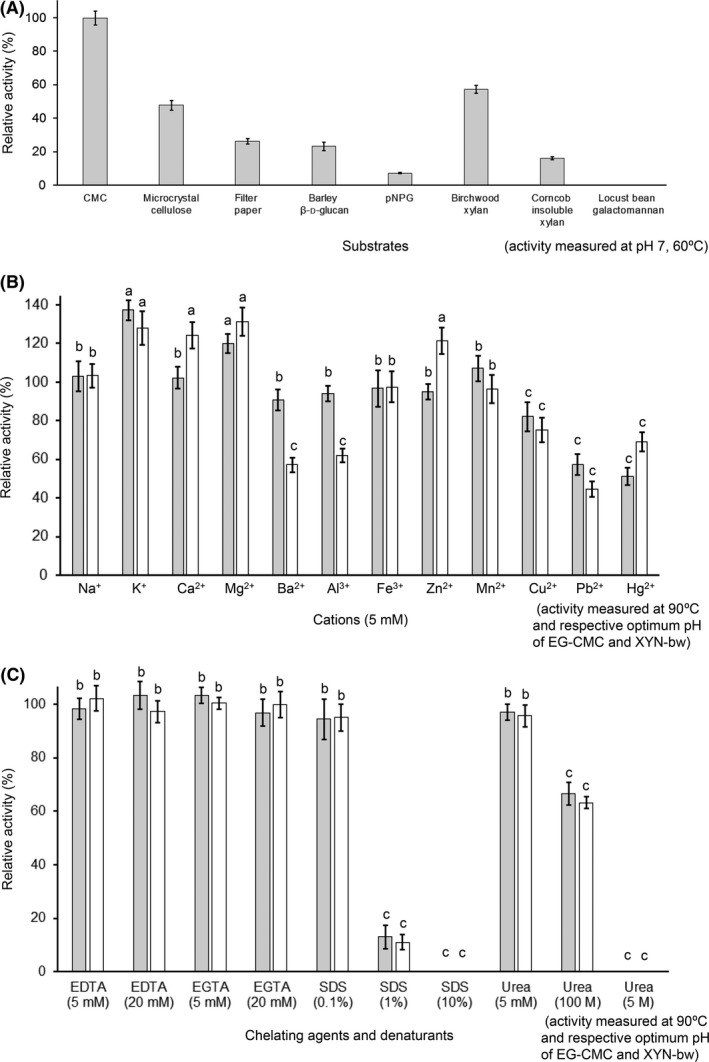
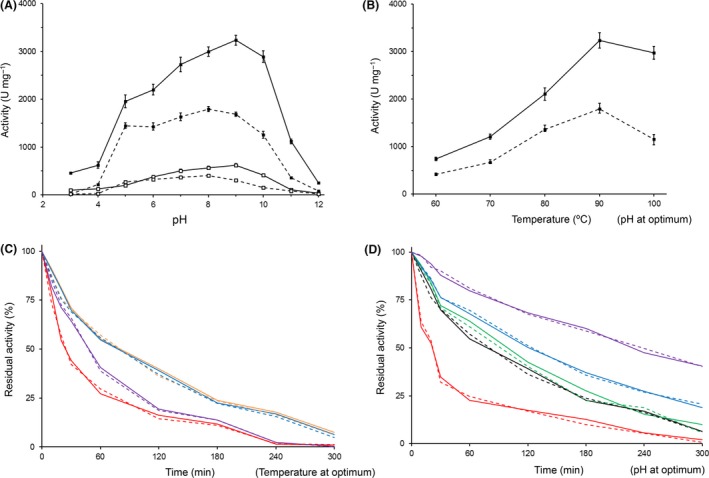
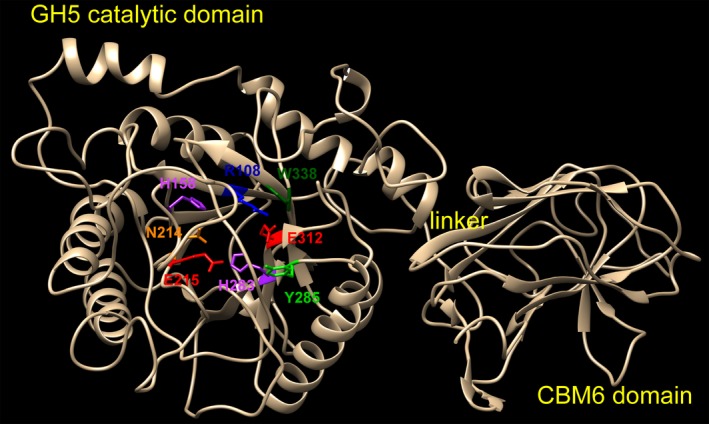
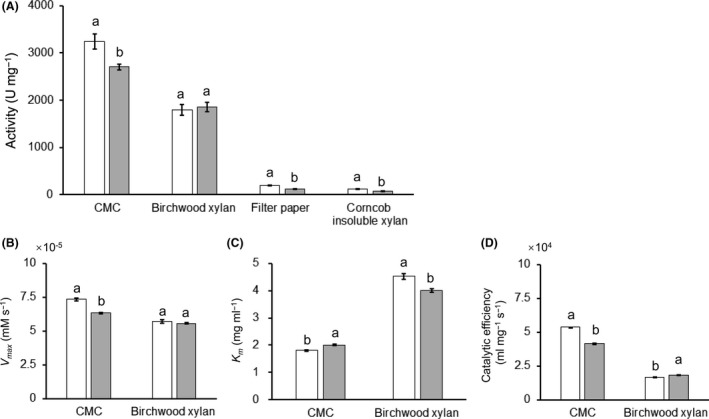
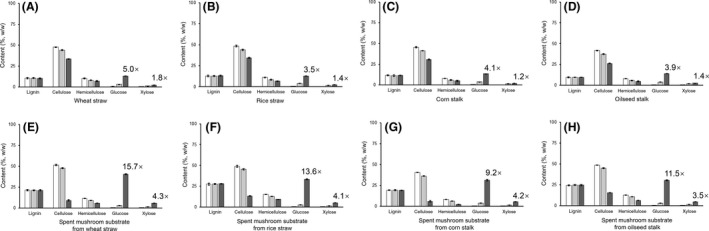
A new cellulolytic strain of Chryseobacterium genus was screened from the dung of a cattle fed with cereal straw. A putative cellulase gene (cbGH5) belonging to glycoside hydrolase family 5 subfamily 46 (GH5_46) was identified and cloned by degenerate PCR plus genome walking. The CbGH5 protein was overexpressed in Pichia pastoris, purified and characterized. It is the first bifunctional cellulase-xylanase reported in GH5_46 as well as in Chryseobacterium genus. The enzyme showed an endoglucanase activity on carboxymethylcellulose of 3237 μmol min-1 mg-1 at pH 9, 90 °C and a xylanase activity on birchwood xylan of 1793 μmol min-1 mg-1 at pH 8, 90 °C. The activity level and thermophilicity are in the front rank of all the known cellulases and xylanases. Core hydrophobicity had a positive effect on the thermophilicity of this enzyme. When similar quantity of enzymatic activity units was applied on the straws of wheat, rice, corn and oilseed rape, CbGH5 could obtain 3.5-5.0× glucose and 1.2-1.8× xylose than a mixed commercial cellulase plus xylanase of Novozymes. When applied on spent mushroom substrates made from the four straws, CbGH5 could obtain 9.2-15.7× glucose and 3.5-4.3× xylose than the mixed Novozymes cellulase+xylanase. The results suggest that CbGH5 could be a promising candidate for industrial lignocellulosic biomass conversion.
© 2018 The Authors. Microbial Biotechnology published by John Wiley & Sons Ltd and Society for Applied Microbiology.
Figures
Similar articles
-
Characterization of a bifunctional xylanase/endoglucanase from yak rumen microorganisms.Appl Microbiol Biotechnol. 2011 Jun;90(6):1933-42. doi: 10.1007/s00253-011-3182-x. Epub 2011 Apr 1. Appl Microbiol Biotechnol. 2011. PMID: 21455595
-
Putative endoglucanase PcGH5 from Phanerochaete chrysosporium is a β-xylosidase that cleaves xylans in synergistic action with endo-xylanase.J Biosci Bioeng. 2015 Apr;119(4):416-20. doi: 10.1016/j.jbiosc.2014.09.012. Epub 2014 Oct 7. J Biosci Bioeng. 2015. PMID: 25300189
-
Isolation and characterization of a non-specific endoglucanase from a metagenomic library of goat rumen.World J Microbiol Biotechnol. 2016 Jan;32(1):12. doi: 10.1007/s11274-015-1957-4. Epub 2015 Dec 28. World J Microbiol Biotechnol. 2016. PMID: 26712627
-
Streptomyces thermocerradoensis I3 secretes a novel bifunctional xylanase/endoglucanase under solid-state fermentation.Biotechnol Prog. 2020 Mar;36(2):e2934. doi: 10.1002/btpr.2934. Epub 2019 Dec 13. Biotechnol Prog. 2020. PMID: 31642208
-
Cellulase-free xylanases from Bacillus and other microorganisms.FEMS Microbiol Lett. 2000 Feb 1;183(1):1-7. doi: 10.1111/j.1574-6968.2000.tb08925.x. FEMS Microbiol Lett. 2000. PMID: 10650194 Review.
Cited by
-
Extremophiles, a Nifty Tool to Face Environmental Pollution: From Exploitation of Metabolism to Genome Engineering.Int J Environ Res Public Health. 2021 May 14;18(10):5228. doi: 10.3390/ijerph18105228. Int J Environ Res Public Health. 2021. PMID: 34069056 Free PMC article. Review.
-
Build Your Own Mushroom Soil: Microbiota Succession and Nutritional Accumulation in Semi-Synthetic Substratum Drive the Fructification of a Soil-Saprotrophic Morel.Front Microbiol. 2021 May 24;12:656656. doi: 10.3389/fmicb.2021.656656. eCollection 2021. Front Microbiol. 2021. PMID: 34108948 Free PMC article.
-
Characterization of a novel bifunctional enzyme from buffalo rumen metagenome and its effect on in vitro ruminal fermentation and microbial community composition.Anim Nutr. 2023 Jan 14;13:137-149. doi: 10.1016/j.aninu.2023.01.004. eCollection 2023 Jun. Anim Nutr. 2023. PMID: 37123618 Free PMC article.
-
Biochemical characterization of a key laccase-like multicopper oxidase of artificially cultivable Morchella importuna provides insights into plant-litter decomposition.3 Biotech. 2019 May;9(5):171. doi: 10.1007/s13205-019-1688-6. Epub 2019 Apr 9. 3 Biotech. 2019. PMID: 30997308 Free PMC article.
-
A novel thermostable GH10 xylanase with activities on a wide variety of cellulosic substrates from a xylanolytic Bacillus strain exhibiting significant synergy with commercial Celluclast 1.5 L in pretreated corn stover hydrolysis.Biotechnol Biofuels. 2019 Mar 9;12:48. doi: 10.1186/s13068-019-1389-8. eCollection 2019. Biotechnol Biofuels. 2019. PMID: 30899328 Free PMC article.
References
-
- Assareh, R. , Shahbani Zahiri, H. , Akbari Noghabi, K. , Aminzadeh, S. and Bakhshi khaniki, G. (2012) Characterization of the newly isolated Geobacillus sp. T1, the efficient cellulase‐producer on untreated barley and wheat straws. Bioresour Technol 120, 99–105. - PubMed
-
- Bischoff, K.M. , Rooney, A.P. , Li, X.‐L. , Liu, S. , and Hughes, S.R. (2006) Purification and characterization of a family 5 endoglucanase from a moderately thermophilic strain of Bacillus licheniformis . Biotechnol Lett 28: 1761–1765. - PubMed
-
- Borges, T.A. , Souza, A.T.d. , Squina, F.M. , Riaño‐Pachón, D.M. , Santos, R.A.C.d. , Machado, E. , et al (2014) Biochemical characterization of an endoxylanase from Pseudozyma brasiliensis sp. nov. strain GHG001 isolated from the intestinal tract of Chrysomelidae larvae associated to sugarcane roots. Process Biochem 49, 77–83.
-
- Bradford, M.M. (1976) A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein‐dye binding. Anal Biochem 72: 248–254. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
