

Alternative Oxidase Isoforms Are Differentially Activated by Tricarboxylic Acid Cycle Intermediates
- PMID: 29208641
- PMCID: PMC5813554
- DOI: 10.1104/pp.17.01331
Alternative Oxidase Isoforms Are Differentially Activated by Tricarboxylic Acid Cycle Intermediates
Abstract
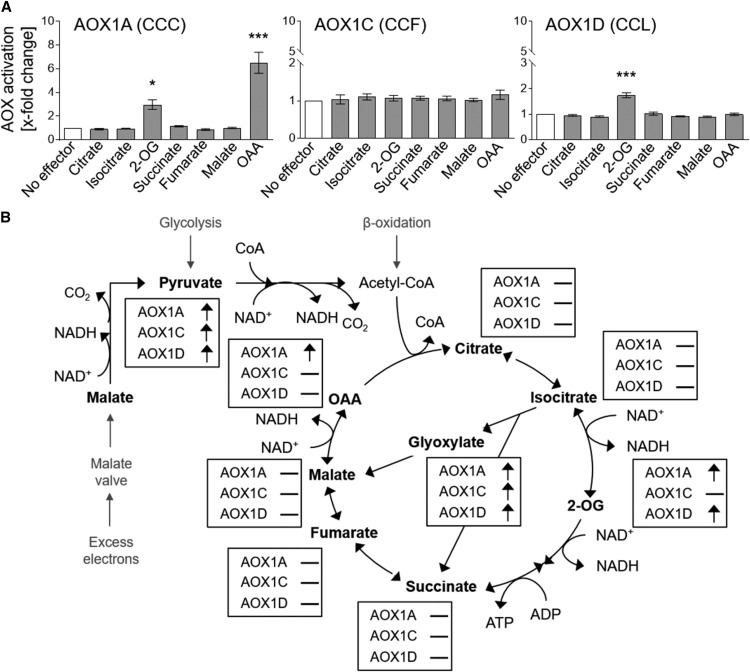
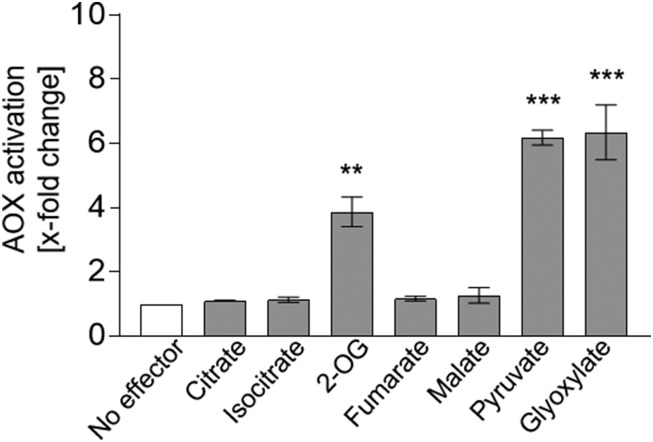
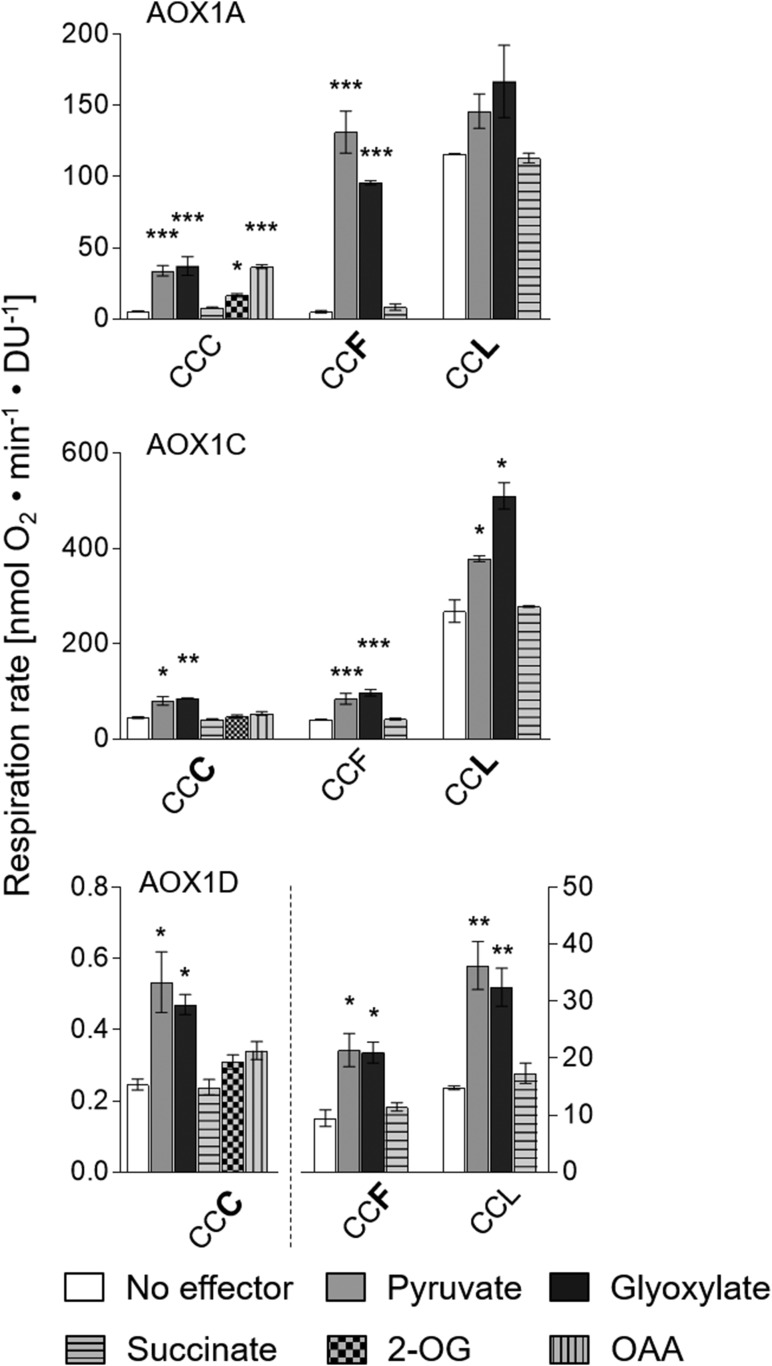
The cyanide-insensitive alternative oxidase (AOX) is a non-proton-pumping ubiquinol oxidase that catalyzes the reduction of oxygen to water and is posttranslationally regulated by redox mechanisms and 2-oxo acids. Arabidopsis (Arabidopsis thaliana) possesses five AOX isoforms (AOX1A-AOX1D and AOX2). AOX1D expression is increased in aox1a knockout mutants from Arabidopsis (especially after restriction of the cytochrome c pathway) but cannot compensate for the lack of AOX1A, suggesting a difference in the regulation of these isoforms. Therefore, we analyzed the different AOX isoenzymes with the aim to identify differences in their posttranslational regulation. Seven tricarboxylic acid cycle intermediates (citrate, isocitrate, 2-oxoglutarate, succinate, fumarate, malate, and oxaloacetate) were tested for their influence on AOX1A, AOX1C, and AOX1D wild-type protein activity using a refined in vitro system. AOX1C is insensitive to all seven organic acids, AOX1A and AOX1D are both activated by 2-oxoglutarate, but only AOX1A is additionally activated by oxaloacetate. Furthermore, AOX isoforms cannot be transformed to mimic one another by substituting the variable cysteine residues at position III in the protein. In summary, we show that AOX isoforms from Arabidopsis are differentially fine-regulated by tricarboxylic acid cycle metabolites (most likely depending on the amino-terminal region around the highly conserved cysteine residues known to be involved in regulation by the 2-oxo acids pyruvate and glyoxylate) and propose that this is the main reason why they cannot functionally compensate for each other.
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Affourtit C, Albury MS, Crichton PG, Moore AL (2002) Exploring the molecular nature of alternative oxidase regulation and catalysis. FEBS Lett 510: 121–126 - PubMed
-
- Alexandre A, Lehninger AL (1984) Bypasses of the antimycin A block of mitochondrial electron transport in relation to ubisemiquinone function. Biochim Biophys Acta 767: 120–129 - PubMed
-
- Bari R, Kebeish R, Kalamajka R, Rademacher T, Peterhänsel C (2004) A glycolate dehydrogenase in the mitochondria of Arabidopsis thaliana. J Exp Bot 55: 623–630 - PubMed
-
- Bouché N, Fromm H (2004) GABA in plants: just a metabolite? Trends Plant Sci 9: 110–115 - PubMed
-
- Campo ML, Kinnally KW, Tedeschi H (1992) The effect of antimycin A on mouse liver inner mitochondrial membrane channel activity. J Biol Chem 267: 8123–8127 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
