

Evolutionarily Conserved nodE, nodO, T1SS, and Hydrogenase System in Rhizobia of Astragalus membranaceus and Caragana intermedia
- PMID: 29209294
- PMCID: PMC5702008
- DOI: 10.3389/fmicb.2017.02282
Evolutionarily Conserved nodE, nodO, T1SS, and Hydrogenase System in Rhizobia of Astragalus membranaceus and Caragana intermedia
Abstract
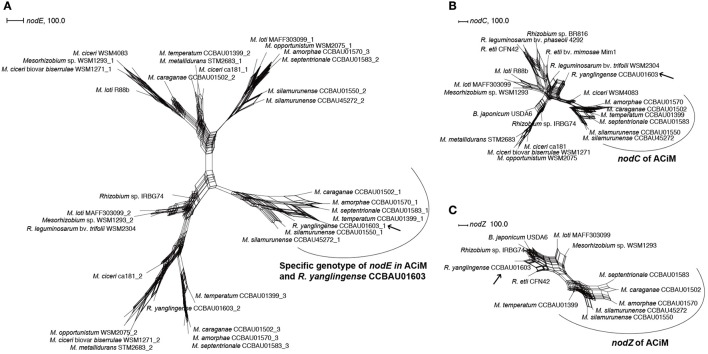
Mesorhizobium species are the main microsymbionts associated with the medicinal or sand-fixation plants Astragalus membranaceus and Caragana intermedia (AC) in temperate regions of China, while all the Mesorhizobium strains isolated from each of these plants could nodulate both of them. However, Rhizobium yanglingense strain CCBAU01603 could nodulate AC plants and it's a high efficiency symbiotic and competitive strain with Caragana. Therefore, the common features shared by these symbiotic rhizobia in genera of Mesorhizobium and Rhizobium still remained undiscovered. In order to study the genomic background influencing the host preference of these AC symbiotic strains, the whole genomes of two (M. silamurunense CCBAU01550, M. silamurunense CCBAU45272) and five representative strains (M. septentrionale CCBAU01583, M. amorphae CCBAU01570, M. caraganae CCBAU01502, M. temperatum CCBAU01399, and R. yanglingense CCBAU01603) originally isolated from AC plants were sequenced, respectively. As results, type III secretion systems (T3SS) of AC rhizobia evolved in an irregular pattern, while an evolutionarily specific region including nodE, nodO, T1SS, and a hydrogenase system was detected to be conserved in all these AC rhizobia. Moreover, nodO was verified to be prevalently distributed in other AC rhizobia and was presumed as a factor affecting the nodule formation process. In conclusion, this research interpreted the multifactorial features of the AC rhizobia that may be associated with their host specificity at cross-nodulation group, including nodE, nodZ, T1SS as the possible main determinants; and nodO, hydrogenase system, and T3SS as factors regulating the bacteroid formation or nitrogen fixation efficiency.
Keywords: Astragalus; Caragana; Mesorhizobium; T1SS; hydrogenase system; nodE; nodO; symbiotic specificity.
Figures






References
-
- Bloemberg G. V., Kamst E., Harteveld M., van Der Drift K. M., Haverkamp J., Thomasoates J. E., et al. (1995). A central domain of Rhizobium NodE protein mediates host specificity by determining the hydrophobicity of fatty acyl moieties of nodulation factors. Mol. Microbiol. 16, 1123–1136. 10.1111/j.1365-2958.1995.tb02337.x - DOI - PubMed
-
- Delcher A. (2006). GlimmerReleaseNotes Version3.02. Available online at: https://ccb.jhu.edu/software/glimmer
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials