

Distribution of cardiac sodium channels in clusters potentiates ephaptic interactions in the intercalated disc
- PMID: 29210458
- PMCID: PMC5813604
- DOI: 10.1113/JP275351
Distribution of cardiac sodium channels in clusters potentiates ephaptic interactions in the intercalated disc
Abstract
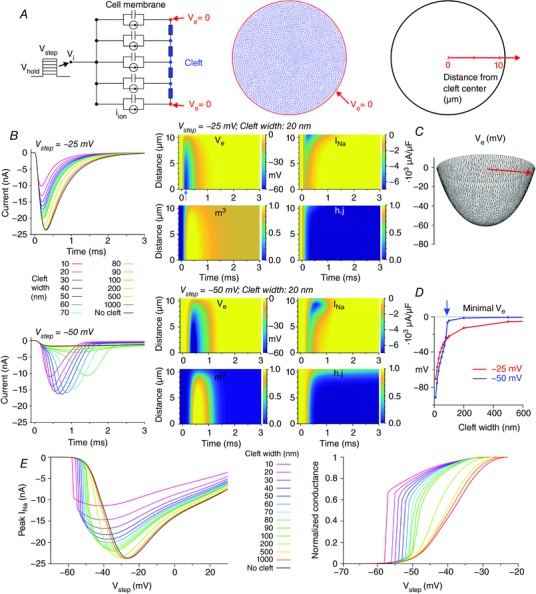
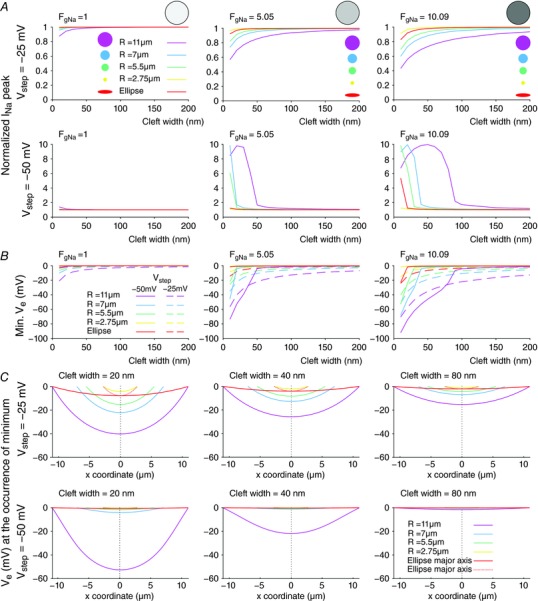
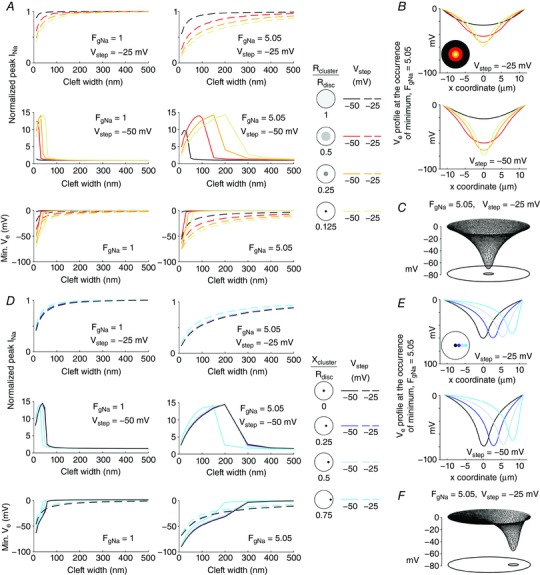
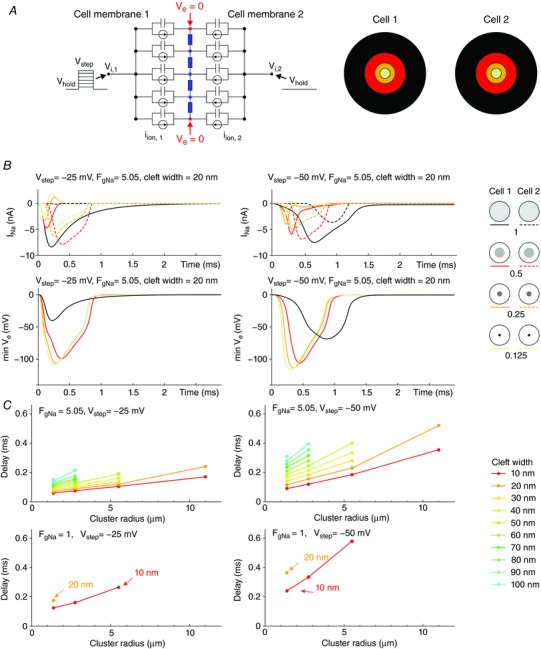
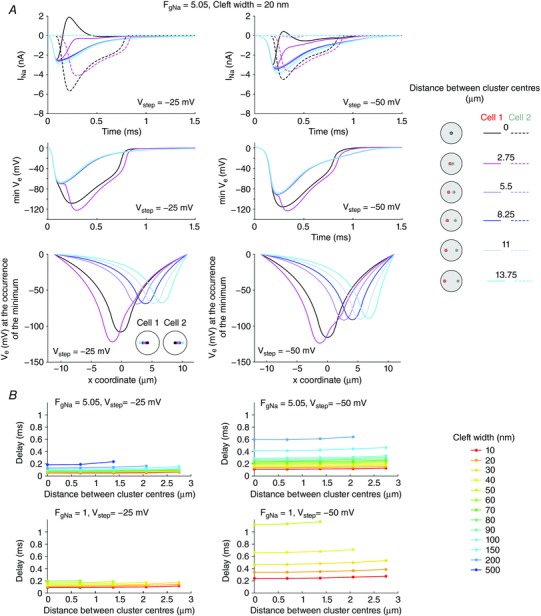
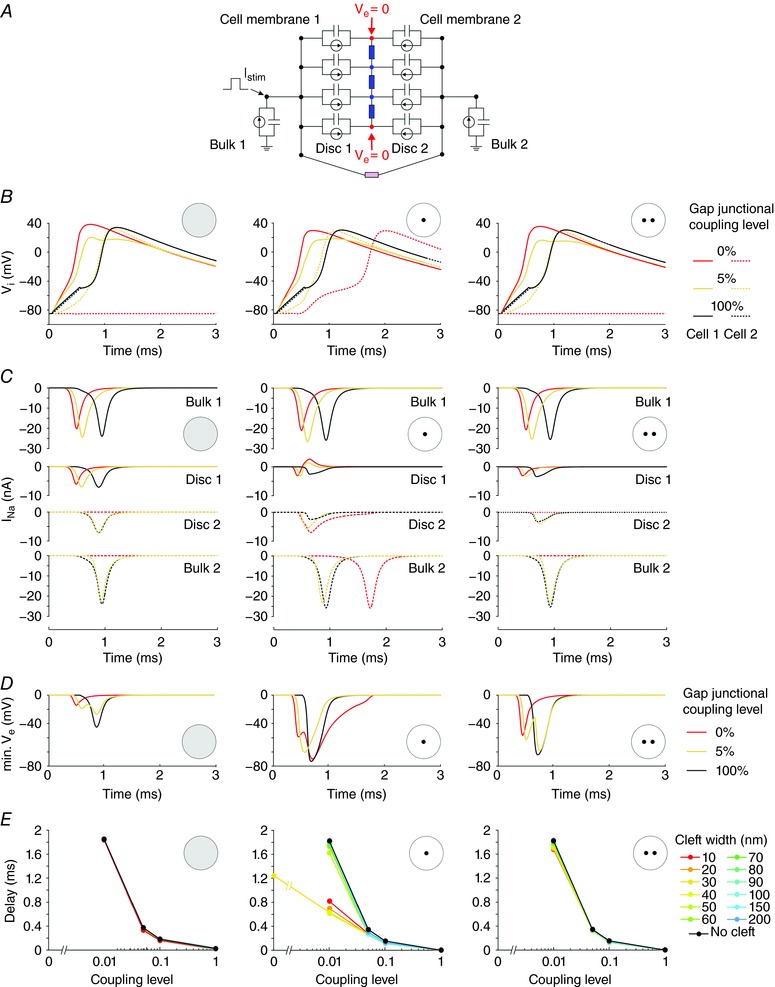
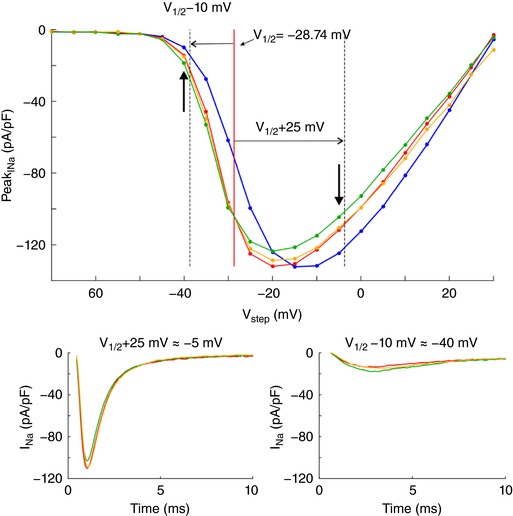
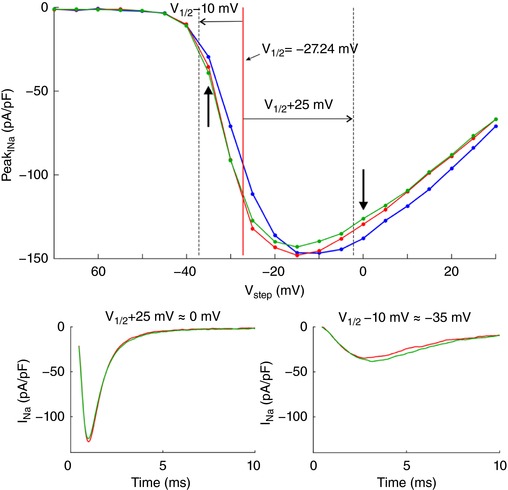
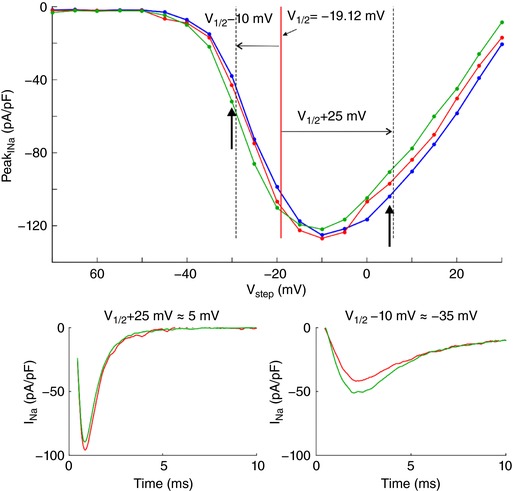
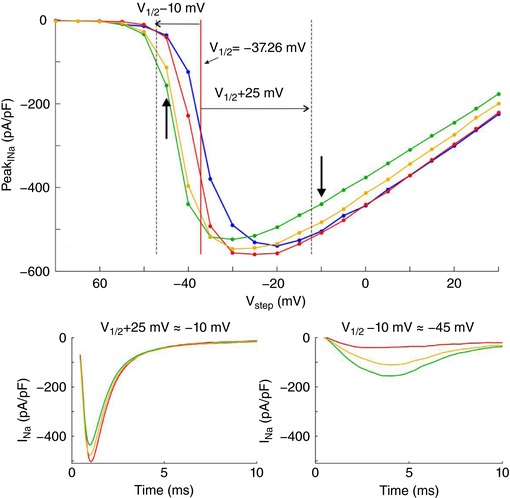
Key points: It has been proposed that ephaptic conduction, relying on interactions between the sodium (Na+ ) current and the extracellular potential in intercalated discs, might contribute to cardiac conduction when gap junctional coupling is reduced, but this mechanism is still controversial. In intercalated discs, Na+ channels form clusters near gap junction plaques, but the functional significance of these clusters has never been evaluated. In HEK cells expressing cardiac Na+ channels, we show that restricting the extracellular space modulates the Na+ current, as predicted by corresponding simulations accounting for ephaptic effects. In a high-resolution model of the intercalated disc, clusters of Na+ channels that face each other across the intercellular cleft facilitate ephaptic impulse transmission when gap junctional coupling is reduced. Thus, our simulations reveal a functional role for the clustering of Na+ channels in intercalated discs, and suggest that rearrangement of these clusters in disease may influence cardiac conduction.
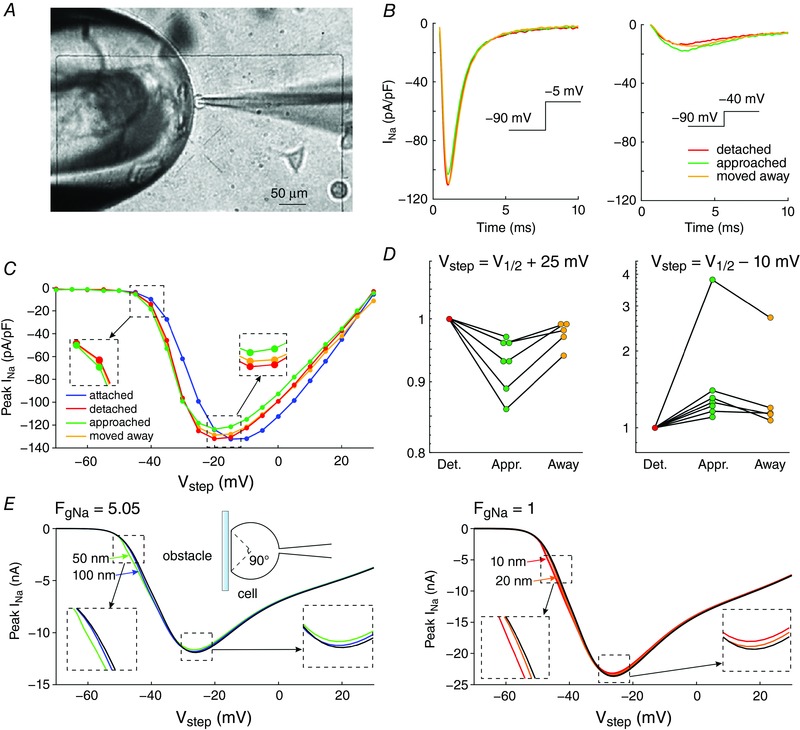
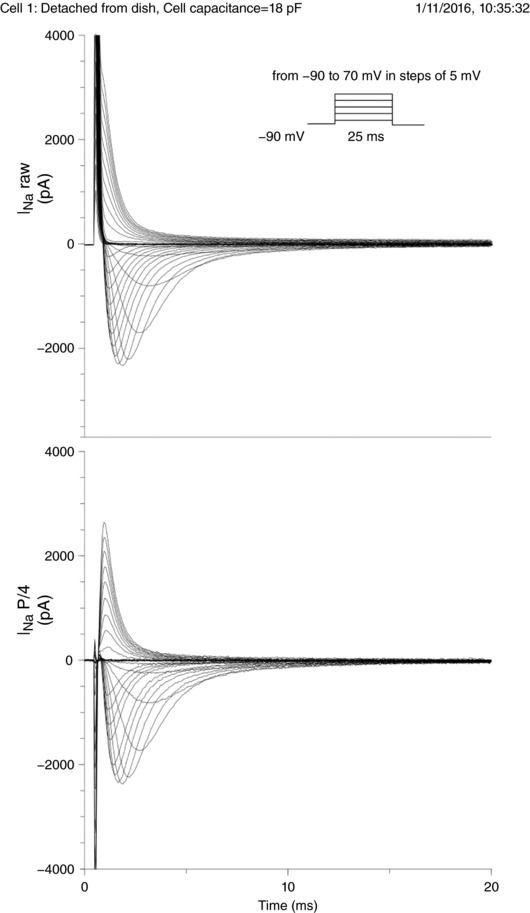
Abstract: It has been proposed that ephaptic interactions in intercalated discs, mediated by extracellular potentials, contribute to cardiac impulse propagation when gap junctional coupling is reduced. However, experiments demonstrating ephaptic effects on the cardiac Na+ current (INa ) are scarce. Furthermore, Na+ channels form clusters around gap junction plaques, but the electrophysiological significance of these clusters has never been investigated. In patch clamp experiments with HEK cells stably expressing human Nav 1.5 channels, we examined how restricting the extracellular space modulates INa elicited by an activation protocol. In parallel, we developed a high-resolution computer model of the intercalated disc to investigate how the distribution of Na+ channels influences ephaptic interactions. Approaching the HEK cells to a non-conducting obstacle always increased peak INa at step potentials near the threshold of INa activation and decreased peak INa at step potentials far above threshold (7 cells, P = 0.0156, Wilcoxon signed rank test). These effects were consistent with corresponding control simulations with a uniform Na+ channel distribution. In the intercalated disc computer model, redistributing the Na+ channels into a central cluster of the disc potentiated ephaptic effects. Moreover, ephaptic impulse transmission from one cell to another was facilitated by clusters of Na+ channels facing each other across the intercellular cleft when gap junctional coupling was reduced. In conclusion, our proof-of-principle experiments demonstrate that confining the extracellular space modulates cardiac INa , and our simulations reveal the functional role of the aggregation of Na+ channels in the perinexus. These findings highlight novel concepts in the physiology of cardiac excitation.
Keywords: action potential; cardiac electrophysiology; computer modelling; ephaptic coupling; intercalated disc; patch clamp; sodium channels; sodium current.
© 2017 The Authors. The Journal of Physiology © 2017 The Physiological Society.
Figures
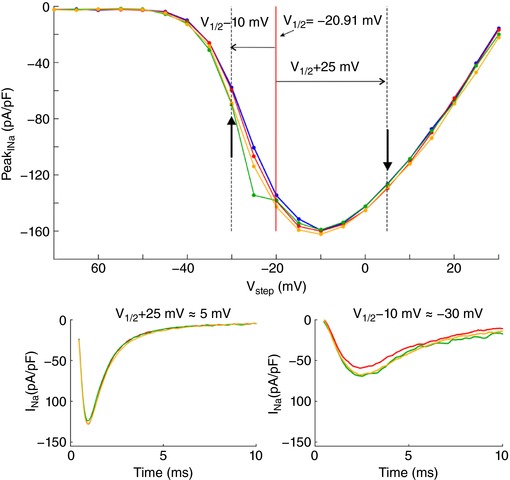
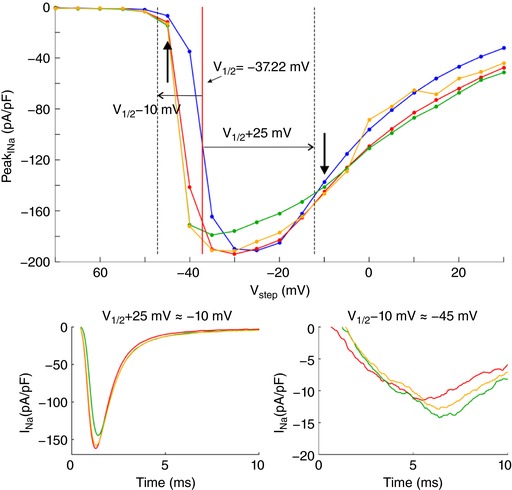
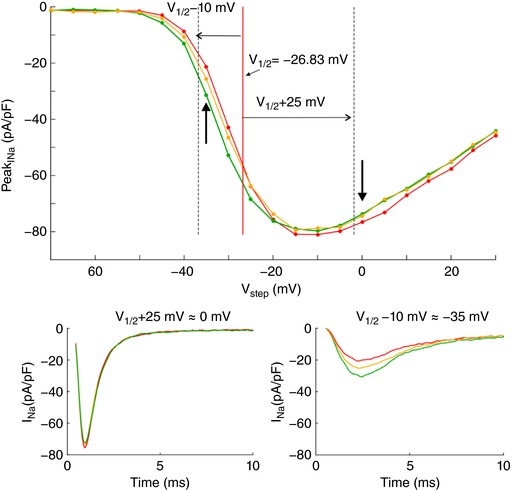
References
-
- Asghari P, Scriven DR, Hoskins J, Fameli N, van Breemen C & Moore ED (2012). The structure and functioning of the couplon in the mammalian cardiomyocyte. Protoplasma 249(Suppl. 1), S31–38. - PubMed
-
- Beauchamp P, Choby C, Desplantez T, de Peyer K, Green K, Yamada KA, Weingart R, Saffitz JE & Kléber AG (2004). Electrical propagation in synthetic ventricular myocyte strands from germline connexin 43 knockout mice. Circ Res 95, 170–178. - PubMed
-
- Cohen SA (1996). Immunocytochemical localization of rH1 sodium channel in adult rat heart atria and ventricle. Presence in terminal intercalated disks. Circulation 94, 3083–3086. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous