

Identification and analysis of Eimeria nieschulzi gametocyte genes reveal splicing events of gam genes and conserved motifs in the wall-forming proteins within the genus Eimeria (Coccidia, Apicomplexa)
- PMID: 29210668
- PMCID: PMC5718062
- DOI: 10.1051/parasite/2017049
Identification and analysis of Eimeria nieschulzi gametocyte genes reveal splicing events of gam genes and conserved motifs in the wall-forming proteins within the genus Eimeria (Coccidia, Apicomplexa)
Abstract
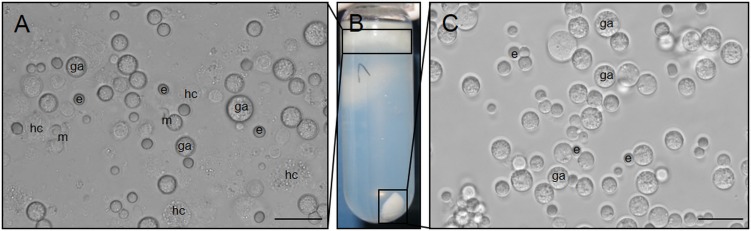
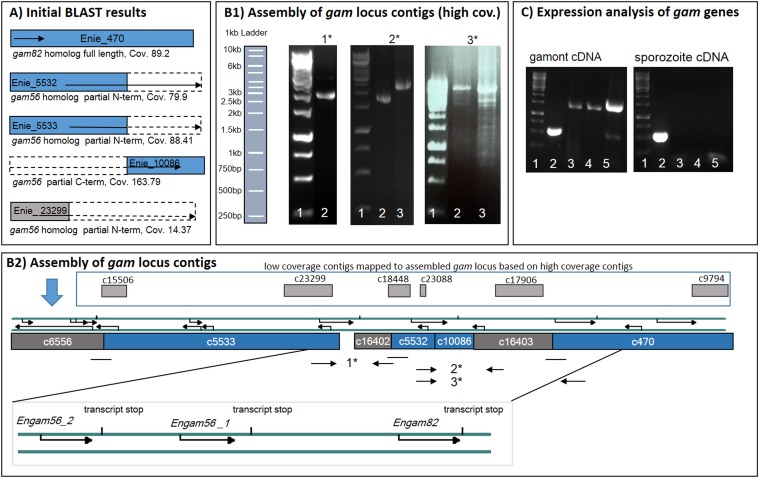
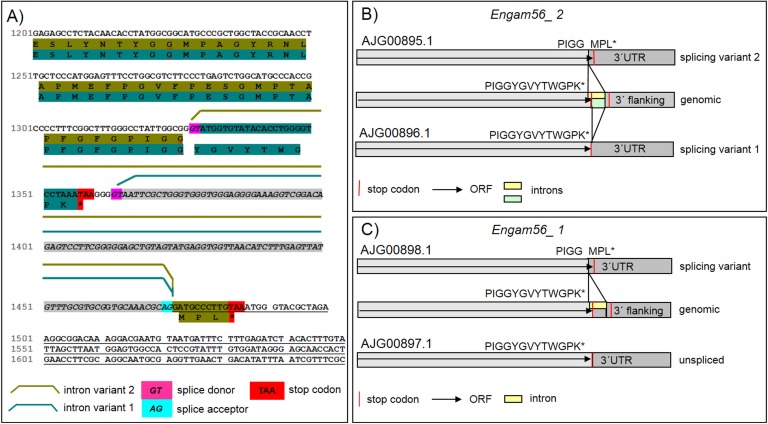
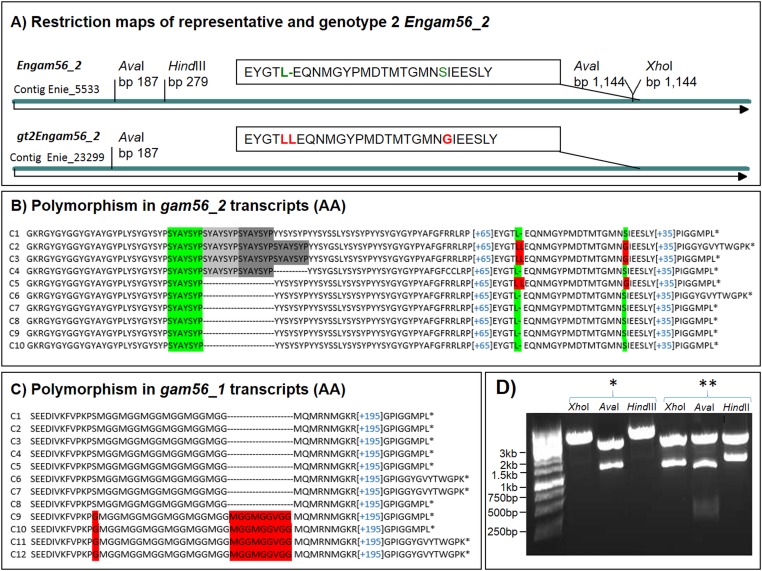
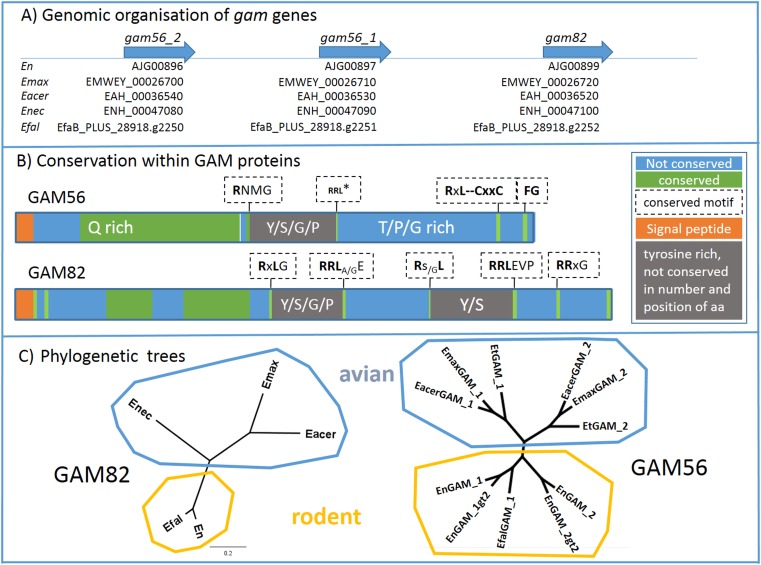
The genus Eimeria (Apicomplexa, Coccidia) provides a wide range of different species with different hosts to study common and variable features within the genus and its species. A common characteristic of all known Eimeria species is the oocyst, the infectious stage where its life cycle starts and ends. In our study, we utilized Eimeria nieschulzi as a model organism. This rat-specific parasite has complex oocyst morphology and can be transfected and even cultivated in vitro up to the oocyst stage. We wanted to elucidate how the known oocyst wall-forming proteins are preserved in this rodent Eimeria species compared to other Eimeria. In newly obtained genomics data, we were able to identify different gametocyte genes that are orthologous to already known gam genes involved in the oocyst wall formation of avian Eimeria species. These genes appeared putatively as single exon genes, but cDNA analysis showed alternative splicing events in the transcripts. The analysis of the translated sequence revealed different conserved motifs but also dissimilar regions in GAM proteins, as well as polymorphic regions. The occurrence of an underrepresented gam56 gene version suggests the existence of a second distinct E. nieschulzi genotype within the E. nieschulzi Landers isolate that we maintain.
Le genre Eimeria (Apicomplexa, Coccidia) fournit une large variété de différentes espèces avec différents hôtes pour étudier les caractéristiques communes et variables dans le genre et ses espèces. Une caractéristique commune de toutes les espèces connues d'Eimeria est l'oocyste, l'étape infectieuse par laquelle son cycle de vie commence et se termine. Dans notre étude, nous avons utilisé Eimeria nieschulzi comme organisme modèle. Ce parasite spécifique au rat présente une morphologie complexe de ses oocystes et peut être transfecté et même cultivé in vitro jusqu'au stade oocyste. Nous voulions élucider comment, dans cette espèce d'Eimeria de rongeurs, les protéines de formation de paroi d'oocystes connues sont préservées, en comparaison aux autres Eimeria. Dans les nouvelles données génomiques obtenues, nous avons pu identifier différents gènes de gamétocytes qui sont orthologues aux gènes gam déjà connus et impliqués dans la formation de la paroi d'oocystes d'espèces d'Eimeria aviaires. Ces gènes apparaissent possiblement comme des exons isolés, mais l'analyse par ADNc a montré des événements d'épissage alternatifs dans les transcripts. L'analyse de la séquence traduite a révélé différents motifs conservés et aussi des régions dissemblables dans les protéines GAM, ainsi que des régions polymorphes. L'existence d'une version sous-représentée du gène gam56 suggère l'existence d'un deuxième génotype distinct d'E. nieschulzi dans l'isolat E. nieschulzi Landers que nous entretenons.
© S. Wiedmer et al., published by EDP Sciences, 2017.
Figures
References
-
- Barta JR, Martin DS, Liberator PA, Dashkevicz M, Anderson JW, Feighner SD, Elbrecht A, Perkins-Barrow A, Jenkins MC, Danforth HD, Ruff MD, Profous-Juchelka H. 1997. Phylogenetic relationships among eight Eimeria species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. Journal of Parasitology, 83(2), 262–271. - PubMed
-
- Bauer C, Dubremetz JF, Entzeroth R. 1995. Characterization of surface antigens of Eimeria nieschulzi (Sporozoa, Eimeriidae) merozoites. Parasitology Research, 81(3), 230-234. - PubMed
-
- Belli SI, Lee M, Thebo P, Wallach MG, Schwartsburd B, Smith NC. 2002. Biochemical characterisation of the 56 and 82 kDa immunodominant gametocyte antigens from Eimeria maxima. International Journal for Parasitology, 32(7), 805-816. - PubMed
-
- Belli SI, Witcombe D, Wallach MG, Smith NC. 2002. Functional genomics of gam56: characterisation of the role of a 56 kilodalton sexual stage antigen in oocyst wall formation in Eimeria maxima. International Journal for Parasitology, 32(14), 1727-1737. - PubMed
-
- Belli SI, Wallach MG, Luxford C, Davies MJ, Smith NC. 2003. Roles of tyrosine-rich precursor glycoproteins and dityrosine- and 3,4-dihydroxyphenylalanine-mediated protein cross-linking in development of the oocyst wall in the coccidian parasite Eimeria maxima. Eukaryotic Cell, 2(3), 456-464. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources