

Runx3 programs CD8+ T cell residency in non-lymphoid tissues and tumours
- PMID: 29211713
- PMCID: PMC5747964
- DOI: 10.1038/nature24993
Runx3 programs CD8+ T cell residency in non-lymphoid tissues and tumours
Erratum in
-
Erratum: Runx3 programs CD8+ T cell residency in non-lymphoid tissues and tumours.Nature. 2018 Feb 15;554(7692):392. doi: 10.1038/nature25445. Epub 2018 Jan 10. Nature. 2018. PMID: 29320476
Abstract
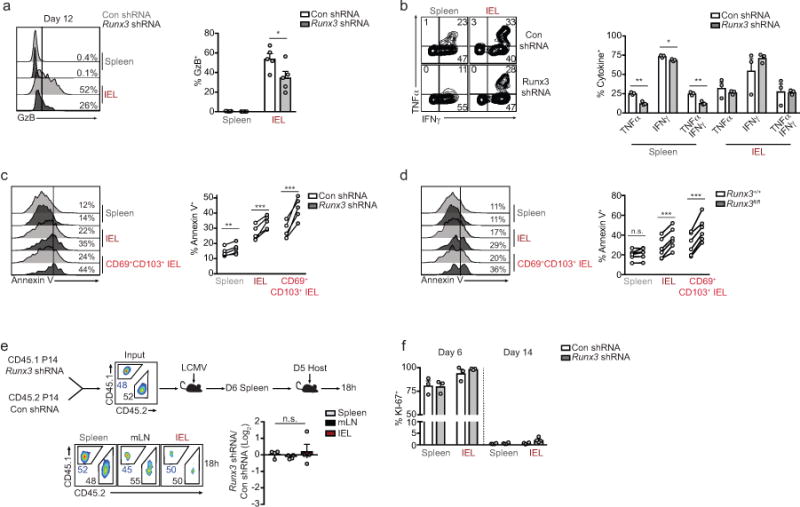
Tissue-resident memory CD8+ T (TRM) cells are found at common sites of pathogen exposure, where they elicit rapid and robust protective immune responses. However, the molecular signals that control TRM cell differentiation and homeostasis are not fully understood. Here we show that mouse TRM precursor cells represent a unique CD8+ T cell subset that is distinct from the precursors of circulating memory cell populations at the levels of gene expression and chromatin accessibility. Using computational and pooled in vivo RNA interference screens, we identify the transcription factor Runx3 as a key regulator of TRM cell differentiation and homeostasis. Runx3 was required to establish TRM cell populations in diverse tissue environments, and supported the expression of crucial tissue-residency genes while suppressing genes associated with tissue egress and recirculation. Furthermore, we show that human and mouse tumour-infiltrating lymphocytes share a core tissue-residency gene-expression signature with TRM cells that is associated with Runx3 activity. In a mouse model of adoptive T cell therapy for melanoma, Runx3-deficient CD8+ tumour-infiltrating lymphocytes failed to accumulate in tumours, resulting in greater rates of tumour growth and mortality. Conversely, overexpression of Runx3 enhanced tumour-specific CD8+ T cell abundance, delayed tumour growth, and prolonged survival. In addition to establishing Runx3 as a central regulator of TRM cell differentiation, these results provide insight into the signals that promote T cell residency in non-lymphoid sites, which could be used to enhance vaccine efficacy or adoptive cell therapy treatments that target cancer.
Conflict of interest statement
Figures












Comment in
-
Resident memory T cells: Runx and hide.Sci Immunol. 2018 Jan 5;3(19):eaar5172. doi: 10.1126/sciimmunol.aar5172. Sci Immunol. 2018. PMID: 29305464
-
Securing front-line T cell defense.Immunol Cell Biol. 2018 Mar;96(3):241-242. doi: 10.1111/imcb.12013. Epub 2018 Feb 10. Immunol Cell Biol. 2018. PMID: 29427338 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
