

Desmoglein 3 Order and Dynamics in Desmosomes Determined by Fluorescence Polarization Microscopy
- PMID: 29212005
- PMCID: PMC5768488
- DOI: 10.1016/j.bpj.2017.09.028
Desmoglein 3 Order and Dynamics in Desmosomes Determined by Fluorescence Polarization Microscopy
Abstract
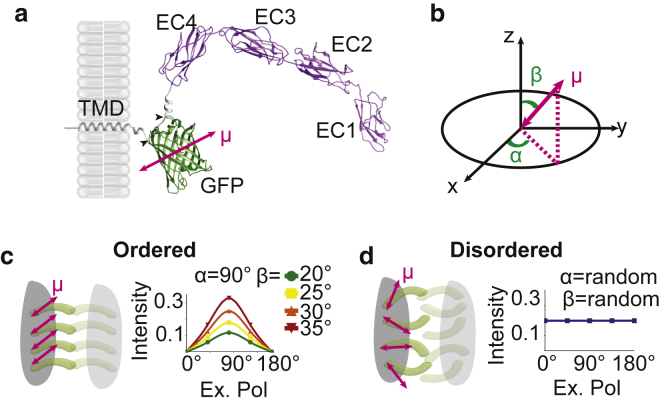
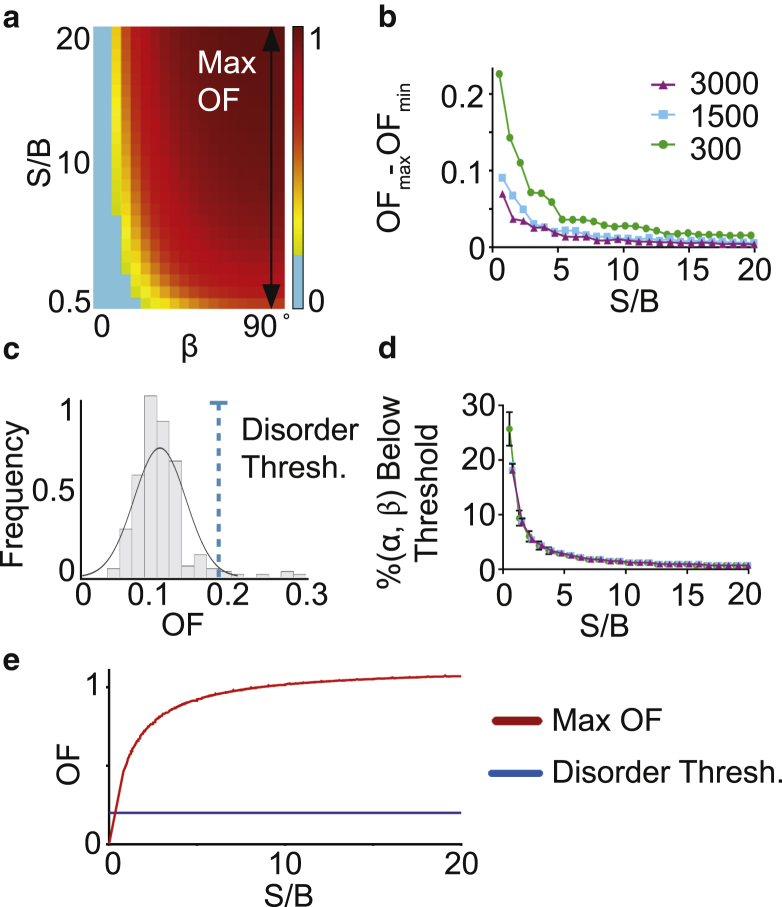
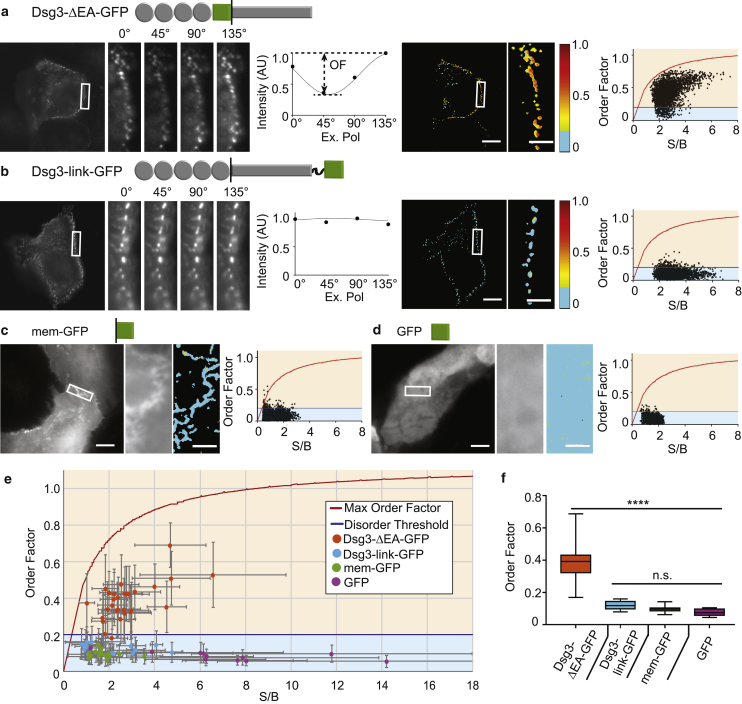
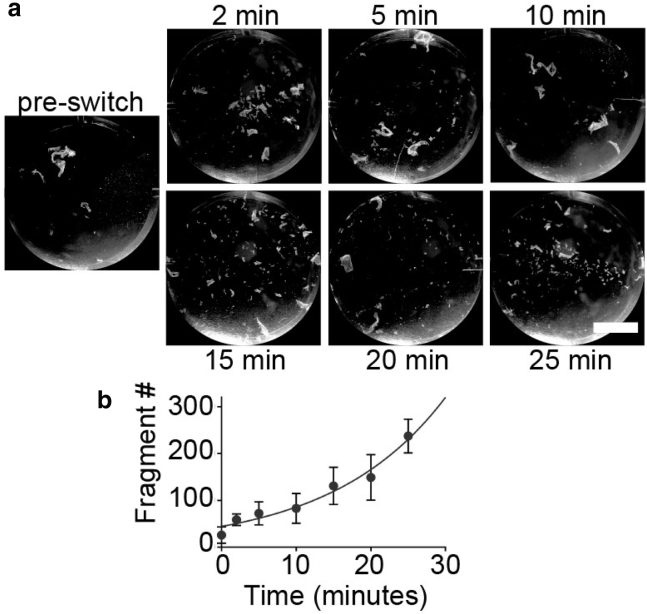
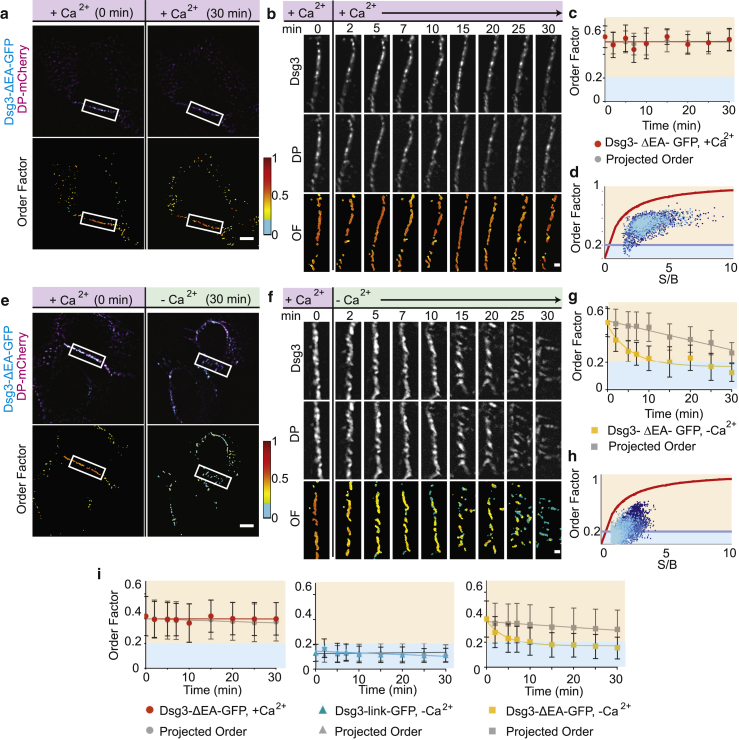
Desmosomes are macromolecular cell-cell junctions that provide adhesive strength in epithelial tissue. Desmosome function is inseparably linked to structure, and it is hypothesized that the arrangement, or order, of desmosomal cadherins in the intercellular space is critical for adhesive strength. However, due to desmosome size, molecular complexity, and dynamics, the role that order plays in adhesion is challenging to study. Herein, we present an excitation resolved fluorescence polarization microscopy approach to measure the spatiotemporal dynamics of order and disorder of the desmosomal cadherin desmoglein 3 (Dsg3) in living cells. Simulations were used to establish order factor as a robust metric for quantifying the spatiotemporal dynamics of order and disorder. Order factor measurements in keratinocytes showed the Dsg3 extracellular domain is ordered at the individual desmosome, single cell, and cell population levels compared to a series of disordered controls. Desmosomal adhesion is Ca2+ dependent, and reduction of extracellular Ca2+ leads to a loss of adhesion measured by dispase fragmentation assay (λ = 15.1 min). Live cell imaging revealed Dsg3 order decreased more rapidly (λ = 5.5 min), indicating that cadherin order is not required for adhesion. Our results suggest that rapid disordering of cadherins can communicate a change in extracellular Ca2+ concentration to the cell, leading to a downstream loss of adhesion. Fluorescence polarization is an effective bridge between protein structure and complex dynamics and the approach presented here is broadly applicable to studying order in macromolecular structures.
Copyright © 2017 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
