

Developmental nonlinearity drives phenotypic robustness
- PMID: 29213092
- PMCID: PMC5719035
- DOI: 10.1038/s41467-017-02037-7
Developmental nonlinearity drives phenotypic robustness
Abstract
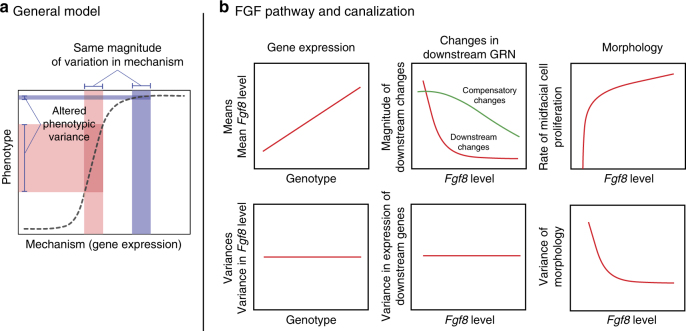
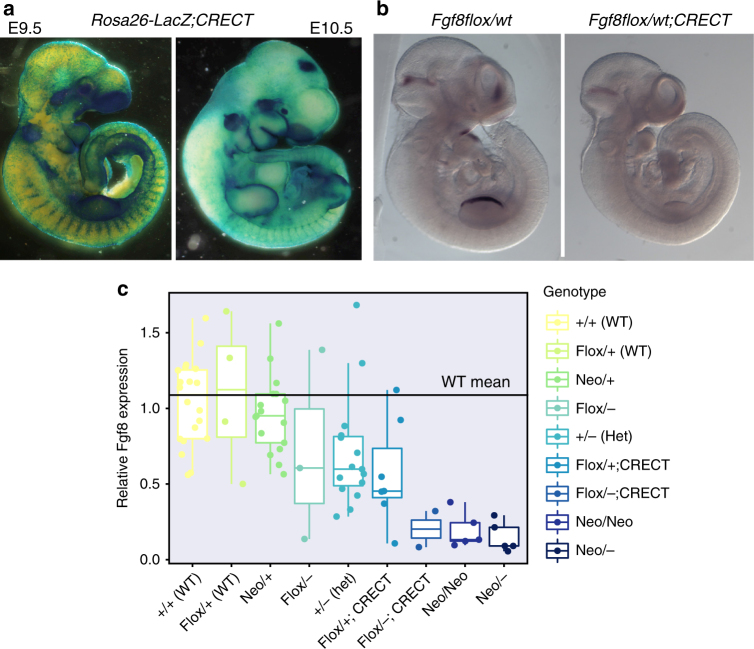
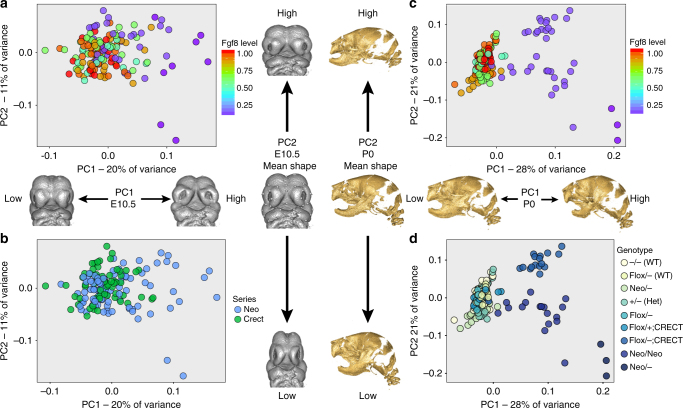
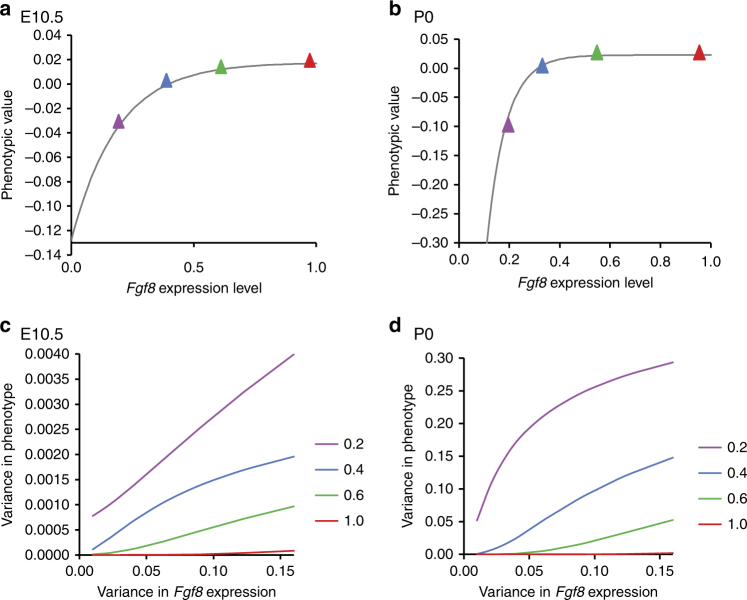
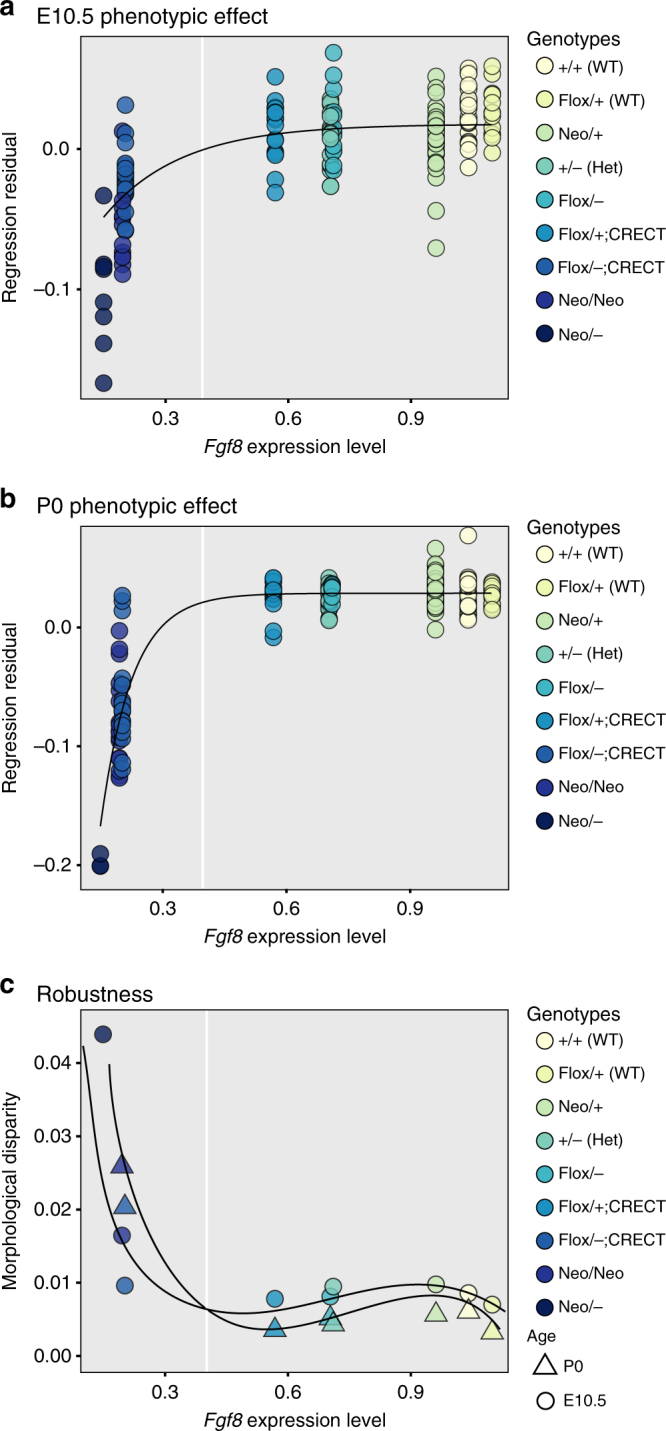
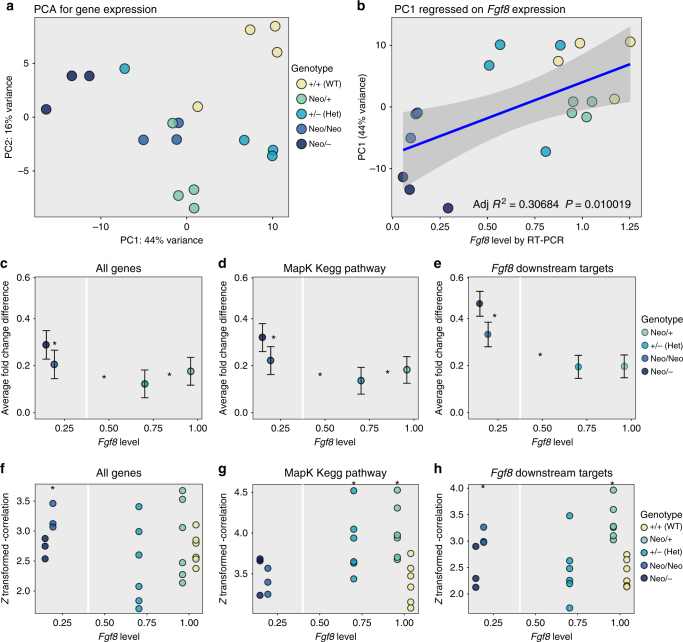
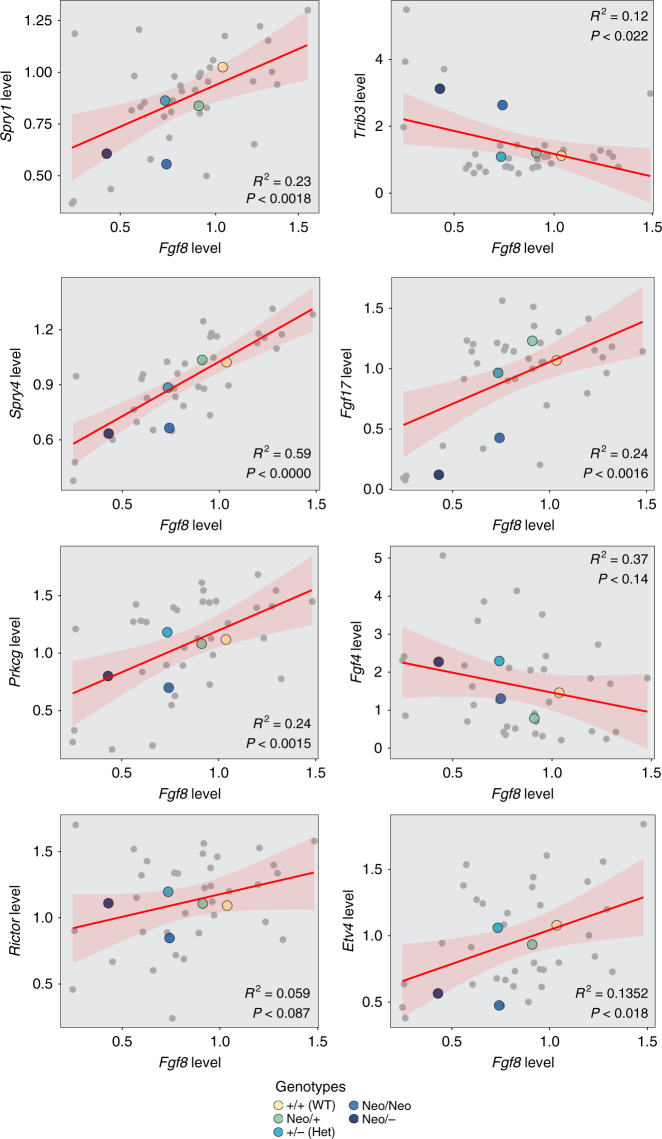
Robustness to perturbation is a fundamental feature of complex organisms. Mutations are the raw material for evolution, yet robustness to their effects is required for species survival. The mechanisms that produce robustness are poorly understood. Nonlinearities are a ubiquitous feature of development that may link variation in development to phenotypic robustness. Here, we manipulate the gene dosage of a signaling molecule, Fgf8, a critical regulator of vertebrate development. We demonstrate that variation in Fgf8 expression has a nonlinear relationship to phenotypic variation, predicting levels of robustness among genotypes. Differences in robustness are not due to gene expression variance or dysregulation, but emerge from the nonlinearity of the genotype-phenotype curve. In this instance, embedded features of development explain robustness differences. How such features vary in natural populations and relate to genetic variation are key questions for unraveling the origin and evolvability of this feature of organismal development.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Waddington CH. The canalisation of development and the inheritance of acquired characters. Nature. 1942;150:563–565. doi: 10.1038/150563a0. - DOI
-
- Waddington CH. Genetic assimilation of the bithorax phenotype. Evolution. 1956;10:1–13. doi: 10.1111/j.1558-5646.1956.tb02824.x. - DOI
-
- Waddington, C. H. The Strategy of the Genes (MacMillan Company, New York, 1957).
-
- Rendel, J. M. Canalization and Gene Control (Logos Press, London, 1967).
-
- Mather K. Genetical control of stability in development. Heredity. 1953;7:297–336. doi: 10.1038/hdy.1953.41. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
