

doi: 10.1038/nsmb.3505.
The ribosome moves: RNA mechanics and translocation
Affiliations
- PMID: 29215639
- PMCID: PMC6581036
- DOI: 10.1038/nsmb.3505
Item in Clipboard
The ribosome moves: RNA mechanics and translocation
Nat Struct Mol Biol.
.
Abstract
During protein synthesis, mRNA and tRNAs must be moved rapidly through the ribosome while maintaining the translational reading frame. This process is coupled to large- and small-scale conformational rearrangements in the ribosome, mainly in its rRNA. The free energy from peptide-bond formation and GTP hydrolysis is probably used to impose directionality on those movements. We propose that the free energy is coupled to two pawls, namely tRNA and EF-G, which enable two ratchet mechanisms to act separately and sequentially on the two ribosomal subunits.
Figures
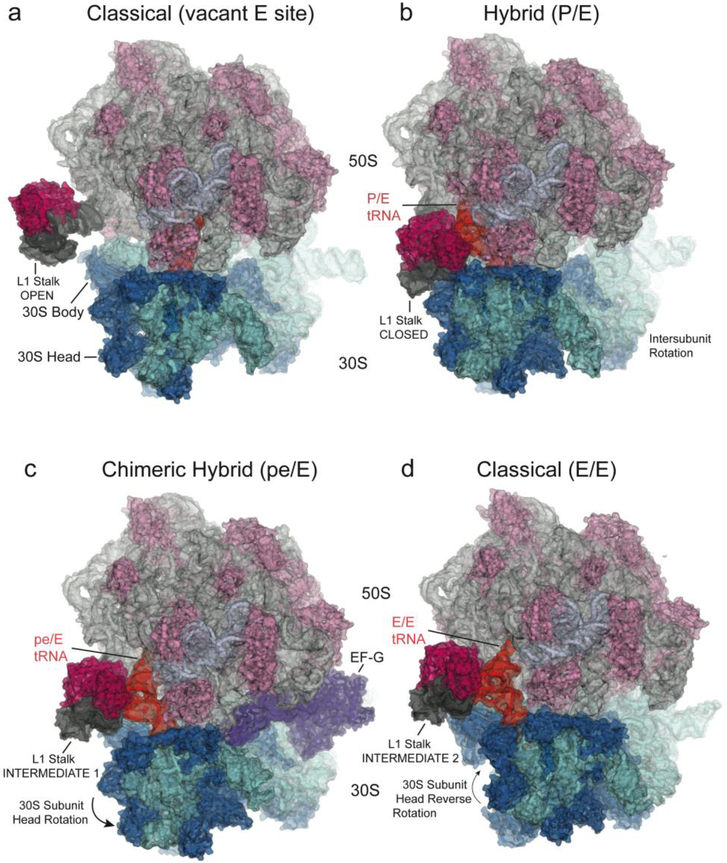
(top) Schematics showing four states of the translocation cycle. (a) Classical (pre-translocation) non-rotated state, with P/P tRNA, vacant E site, and L1 stalk in its open position. (PDB ID: 4GD2)
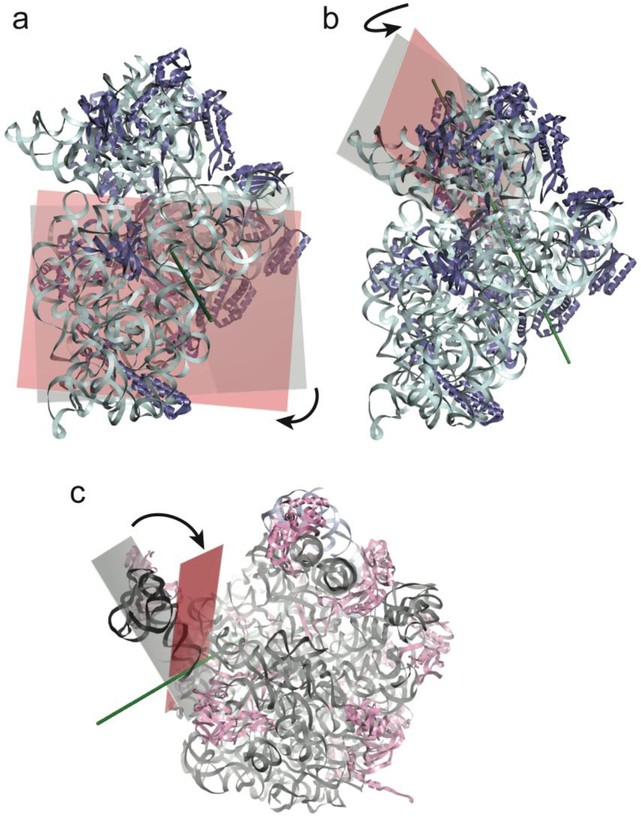
=; (b) Hybrid state, with P/E tRNA, ~6° intersubunit rotation, ~6° 30S subunit head rotation and 30° rotation of the L1 stalk to its closed position, where it contacts the elbow of the P/E tRNA and the 30S subunit. (PDB ID: 4V9H) (c) Chimeric hybrid state, with pe/E tRNA, 21° 30S subunit head rotation, ~2° intersubunit rotation and L1 stalk in Int 1 position. (PDB ID: 4V9K); (d) Classical (post-translocation) non-rotated state, with E/E tRNA, L1 stalk in Int 2 position. (PDB ID: 4V67) Components shown are: 16S rRNA (cyan); 23S rRNA (light grey); 5S rRNA (blue-grey); 30S proteins (dark blue); 50S proteins (light magenta); L1 stalk (dark grey); L1 protein (magenta; docked in panel (a) from Zhou et al. (2013)); deacylated tRNA (orange). The head of the L1 stalk maintains contact with the elbow of the deacylated tRNA as it moves through states b to d. (e-g) The three large-scale motions are shown by planes bisecting the (e) 30S body domain; (f) 30S head domain and (g) L1 stalk in the structures of the non-rotated (cyan or grey) and rotated (pink) states; Euler-Rodrigues axes are represented by grey rods.
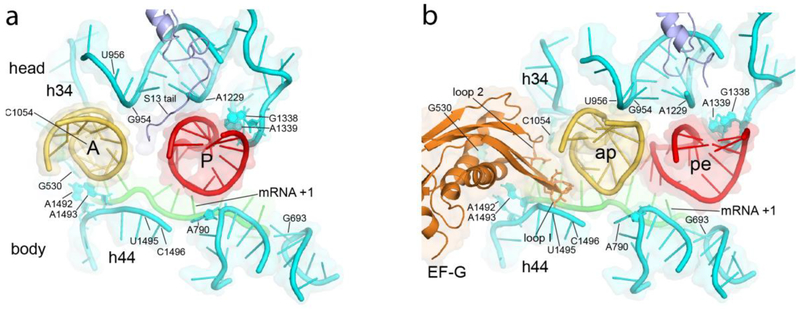
Crystal structures showing positions of the A- and P-site ASLs in the (a) classical-state ribosome, prior to translocation, and in the (b) trapped chimeric hybrid-state translocation intermediate. The P-site ASL moves toward the E site into the pe chimeric state, precisely following rotation of the 30S head domain as shown in (b); the A-site ASL moves further than head rotation, likely following the movement of the tip of domain IV of EF-G into the ap chimeric state. Apart from the tail of protein S13 and domain IV of EF-G, virtually all of the ASL contacts are with 16S ribosomal RNA.
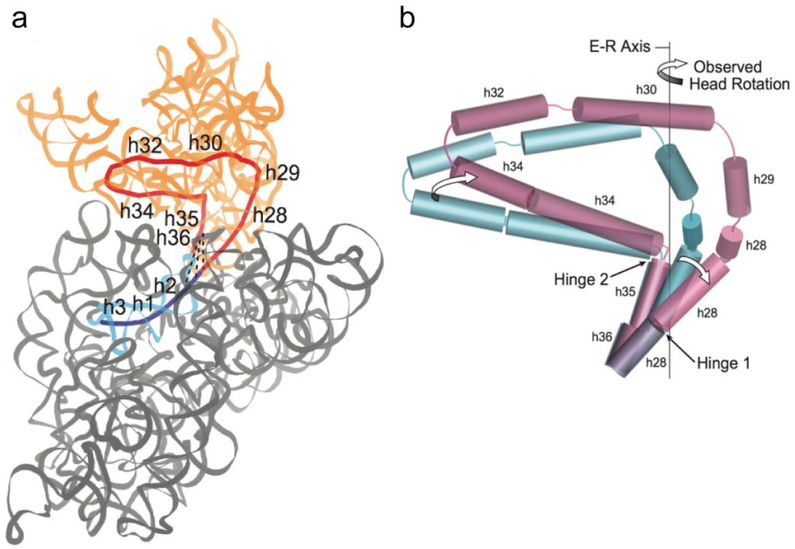
(a) Coaxial helical core of the 16S rRNA component (red) of the 30S head domain (orange) contains helices h28 to h36 (red). Helices h2, h1 and h3 of the body domain (blue) are coaxial with the core helices of the head. Dotted lines represent conserved A-minor interactions between the h36 tetraloop and helix h2 in the 30S body (grey). (b) Positions of coaxial 16S rRNA helical axes are represented by cylinders for the non-rotated (blue) and rotated (magenta) states. During head rotation, movement of Hinge 1 is caused by straightening of the kinked helix h28; movement of Hinge 2 is the result of swiveling of helix h34 around the static h35. Rotation of the 30S head around the Euler-Rodrigues (E-R) axis results from combined bending at Hinges 1 and 2.
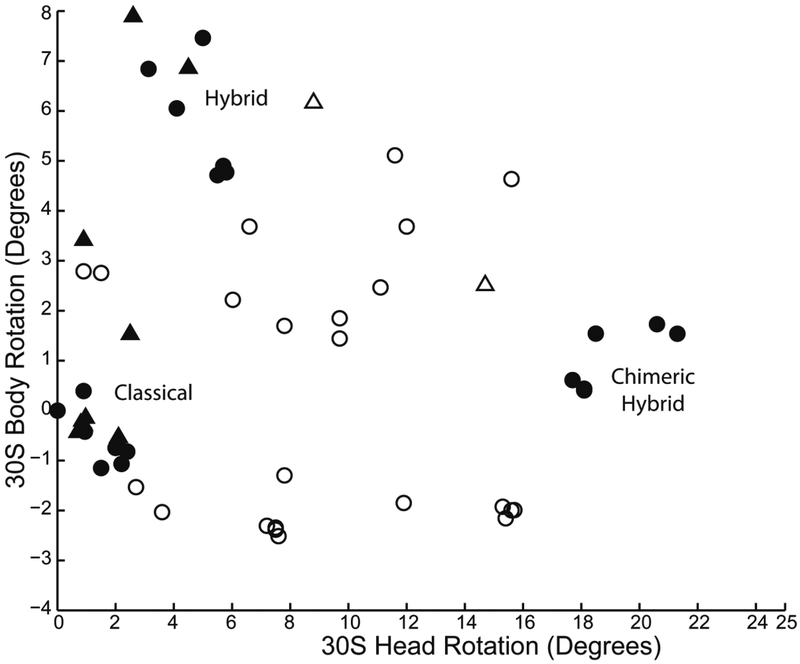
In vacant ribosomes, (empty symbols), head and body rotation angles are distributed across a wide range of possible values. In the presence of tRNA (filled symbols), the ribosomes are dramatically constrained into three clusters, corresponding to the classical, hybrid and chimeric-hybrid states. Data are extracted from 69 X-ray and cryo-EM structures of 8Å or better resolution of bacterial (circles) and eukaryotic (triangles) ribosomes.
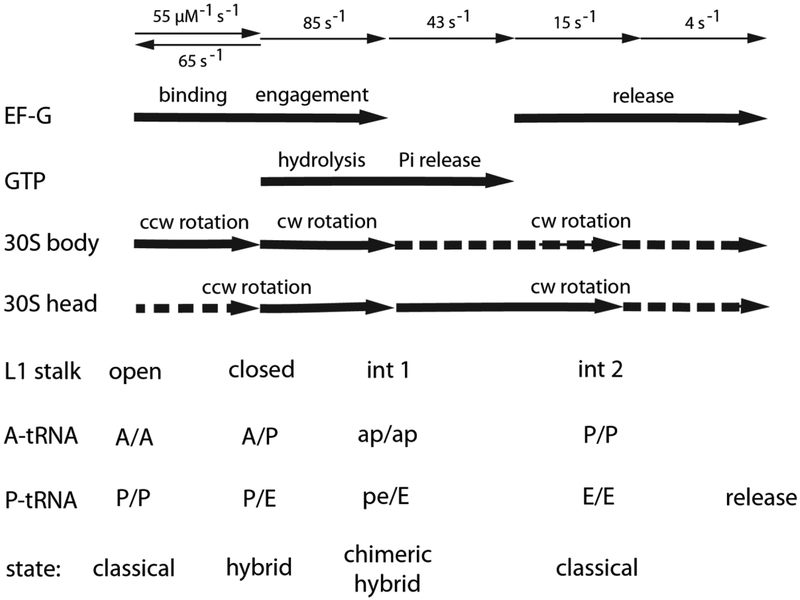
At the top are shown the rates of individual steps of translocation, corresponding to binding, engagement and release of EF-G; and hydrolysis and release of GTP. In the middle are the timing of the corresponding large-scale rotational movements of the 30S body and head domains,,,, and movements of the L1 stalk. At the bottom are the corresponding binding states of the tRNAs,. Dotted arrows indicate smaller-scale rotational movements.
References
-
- Ogle JM et al. Recognition of cognate transfer RNA by the 30S ribosomal subunit. Science 292, 897–902 (2001). - PubMed
-
- Demeshkina N, Jenner L, Westhof E, Yusupov M & Yusupova G A new understanding of the decoding principle on the ribosome. Nature 484, 256–9 (2012). - PubMed
-
- Nissen P, Hansen J, Ban N, Moore PB & Steitz TA The structural basis of ribosome activity in peptide bond synthesis. Science 289, 920–30 (2000). - PubMed
-
- Frank J & Agrawal RK A ratchet-like inter-subunit reorganization of the ribosome during translocation. Nature 406, 318–22 (2000). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
