

Phenotypic characterization of Synechocystis sp. PCC 6803 substrains reveals differences in sensitivity to abiotic stress
- PMID: 29216280
- PMCID: PMC5720811
- DOI: 10.1371/journal.pone.0189130
Phenotypic characterization of Synechocystis sp. PCC 6803 substrains reveals differences in sensitivity to abiotic stress
Abstract
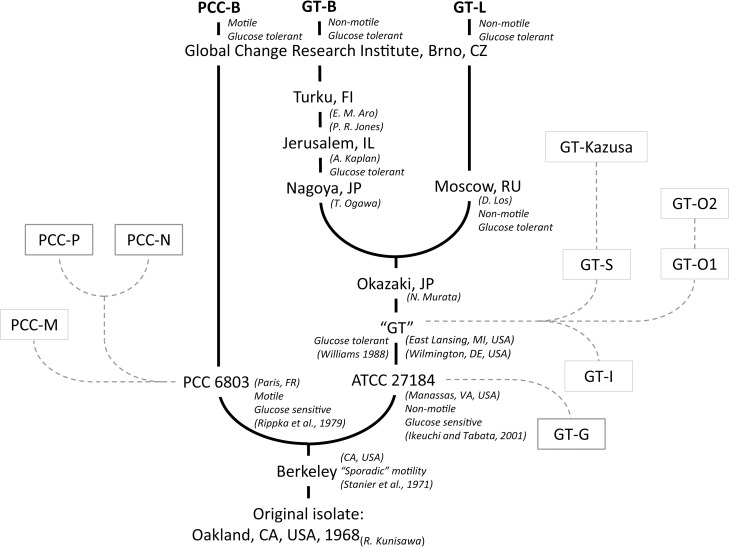
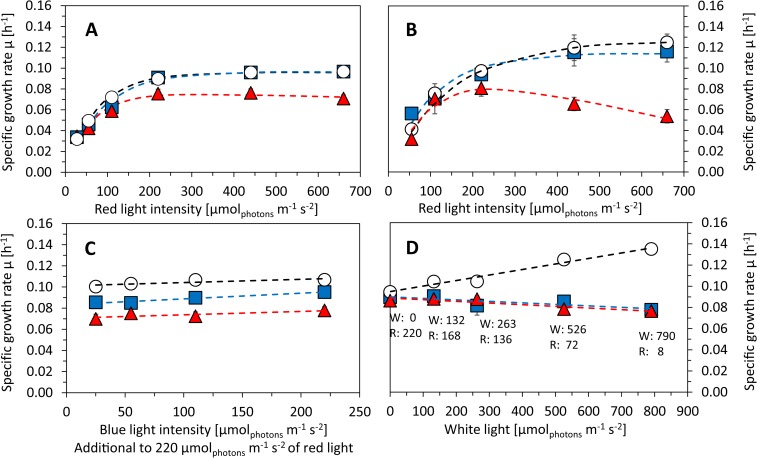
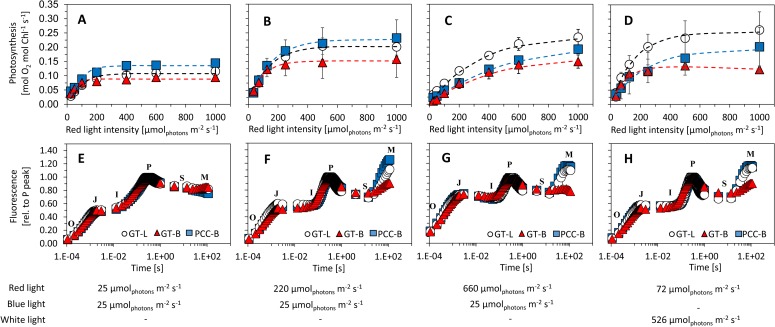
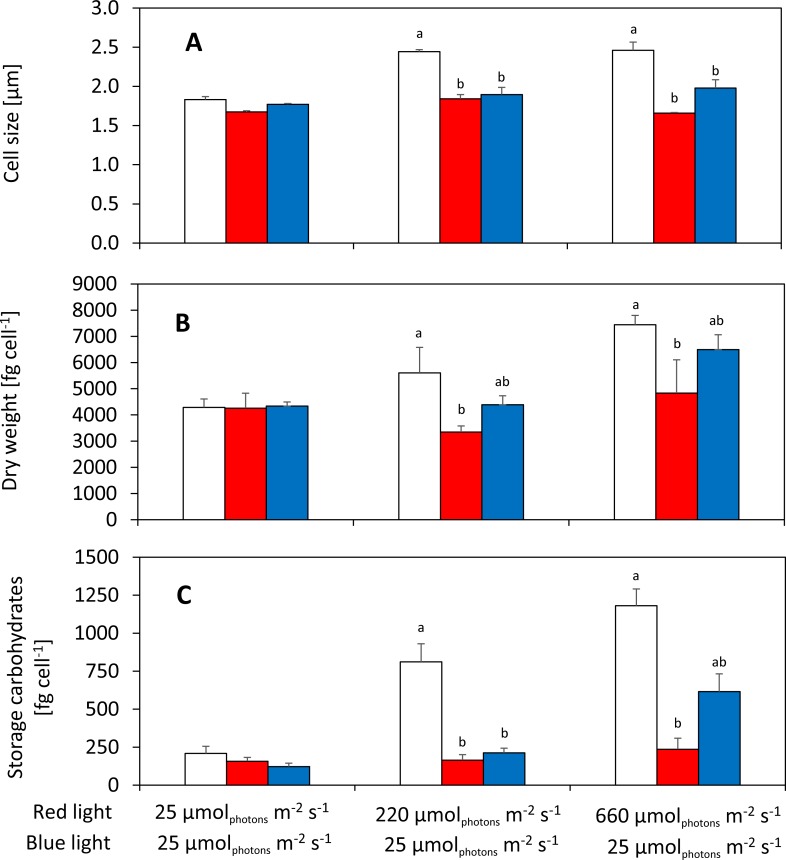
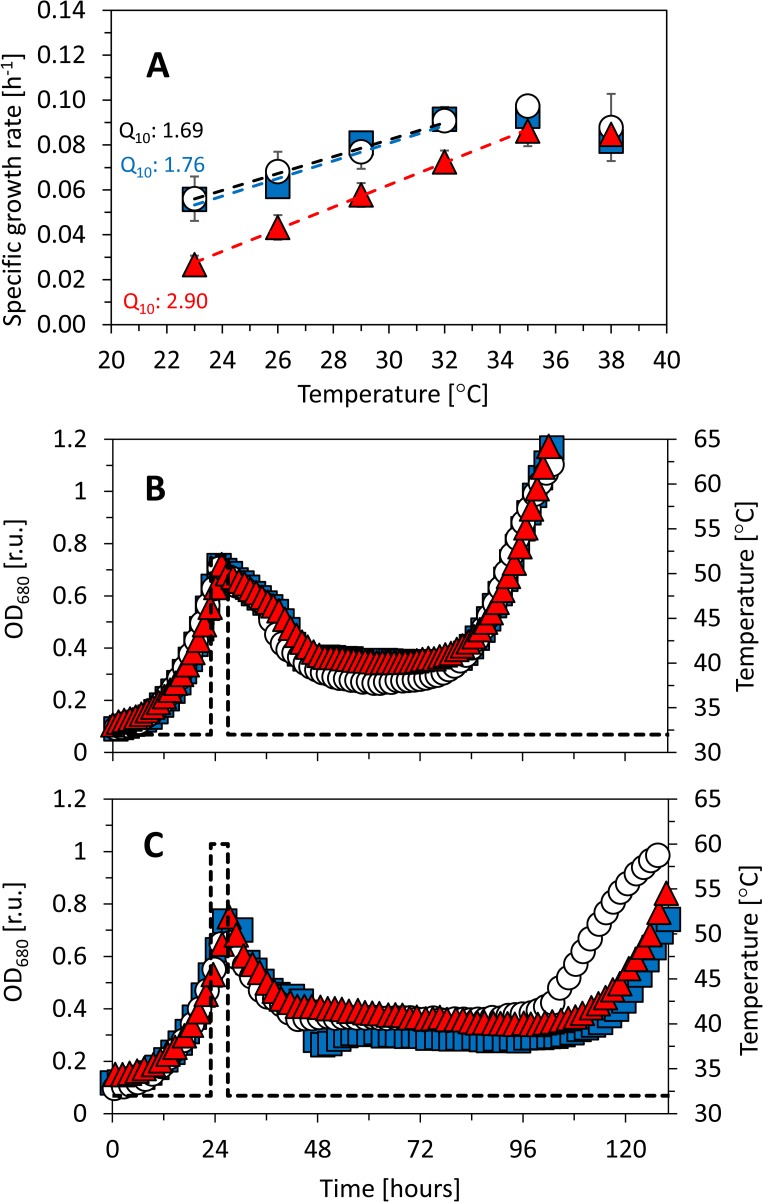
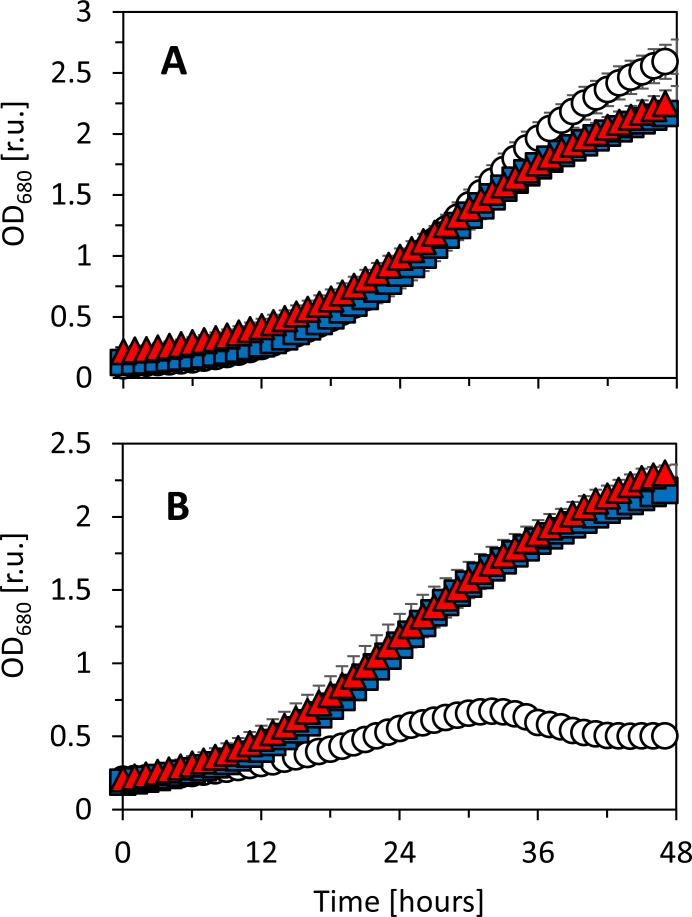
Synechocystis sp. PCC 6803 is a widely used model cyanobacterium, whose substrains can vary on both genotype and phenotype levels. Previously described phenotypic variations include ability of mixotrophic growth, ability of movement on agar plates and variations in pigments composition or cell size. In this study, we report for the first time significant variation among Synechocystis substrains in complex cellular traits such as growth rate, photosynthesis efficiency, cellular dry weight and cellular composition (including protein or carbohydrates content). We also confirmed previously reported differences in cell size. Synechocystis cultures were cultivated in controlled environment of flat panel photobioreactors under red, blue and white light of intensities up to 790 μmol(photons) m-2 s-1, temperatures 23°C-60°C, input CO2 concentrations ranging from 400 to 15 000 ppm and in BG11 cultivation medium with and without addition of NaCl. Three Synechocystis substrains were used for the comparative experiments: GT-L, GT-B (Brno, CZ) and PCC-B (Brno, CZ). Growth rates of Synechocystis GT-B were inhibited under high intensities of red light (585-670 nm), and growth rates of both substrains GT-B and PCC-B were inhibited under photons of wavelengths 485-585 nm and 670-700 nm. Synechocystis GT-B was more sensitive to low temperatures than the other two tested substrains, and Synechocystis GT-L was sensitive to the presence of NaCl in the cultivation media. The results suggest that stress sensitivity of commonly used Synechocystis substrains can strongly vary, similarly as glucose tolerance or motility as reported previously. Our study further supports the previous statement that emphasizes importance of proper Synechocystis substrains selection and awareness of phenotypical differences among Synechocystis substrains which is crucial for comparative and reproducible research. This is highly relevant for studies related to stress physiology and development of sustainable biotechnological applications.
Conflict of interest statement
Figures
References
-
- Sarsekeyeva F, Zayadan BK, Usserbaeva A, Bedbenov VS, Sinetova MA, Los DA. Cyanofuels: Biofuels from cyanobacteria. Reality and perspectives. Photosynth Res. Springer Netherlands; 2015;125: 329–340. doi: 10.1007/s11120-015-0103-3 - DOI - PubMed
-
- Wijffels RH, Kruse O, Hellingwerf KJ. Potential of industrial biotechnology with cyanobacteria and eukaryotic microalgae. Curr Opin Biotechnol. 2013;24: 405–13. doi: 10.1016/j.copbio.2013.04.004 - DOI - PubMed
-
- Kaneko T, Tanaka a, Sato S, Kotani H, Sazuka T, Miyajima N, et al. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. I. Sequence features in the 1 Mb region from map positions 64% to 92% of the genome. DNA Res. 1995;2: 153–166, 191–198. doi: 10.1093/dnares/2.4.153 - DOI - PubMed
-
- Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y, et al. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-coding regions. DNA Res. 1996;3: 109–136. doi: 10.1093/dnares/3.3.109 - DOI - PubMed
-
- Grigorieva G, Shestakov S. Transformation in the cyanobacterium Synechocystis sp. 6803. FEMS Microbiol Lett. 1982;13: 367–370. Available: http://onlinelibrary.wiley.com/doi/10.1111/j.1574-6968.1982.tb08289.x/full - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
