

ApoE4 Accelerates Early Seeding of Amyloid Pathology
- PMID: 29216449
- PMCID: PMC5948105
- DOI: 10.1016/j.neuron.2017.11.013
ApoE4 Accelerates Early Seeding of Amyloid Pathology
Abstract
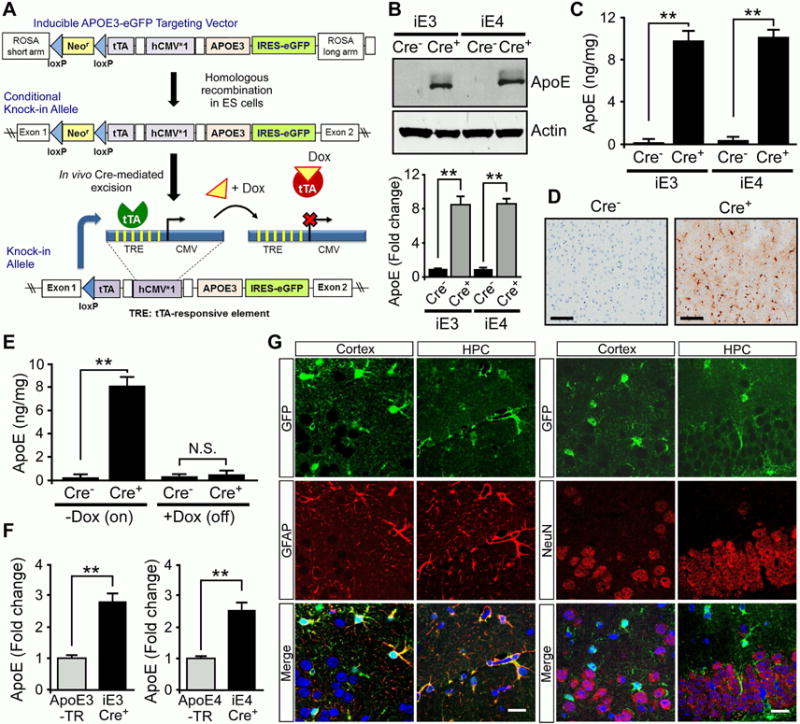
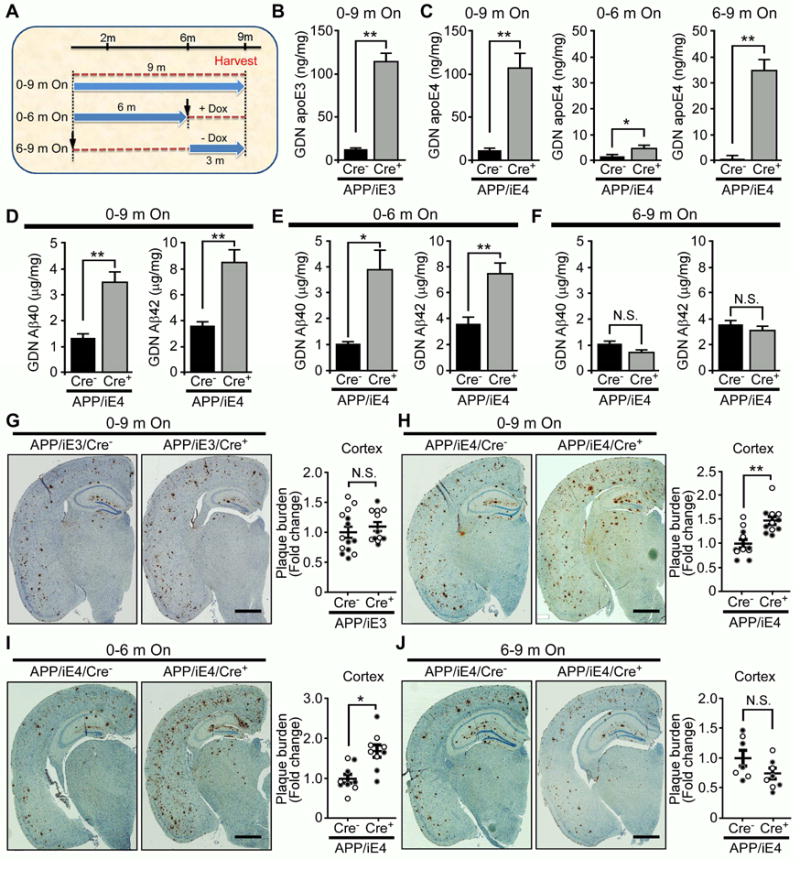
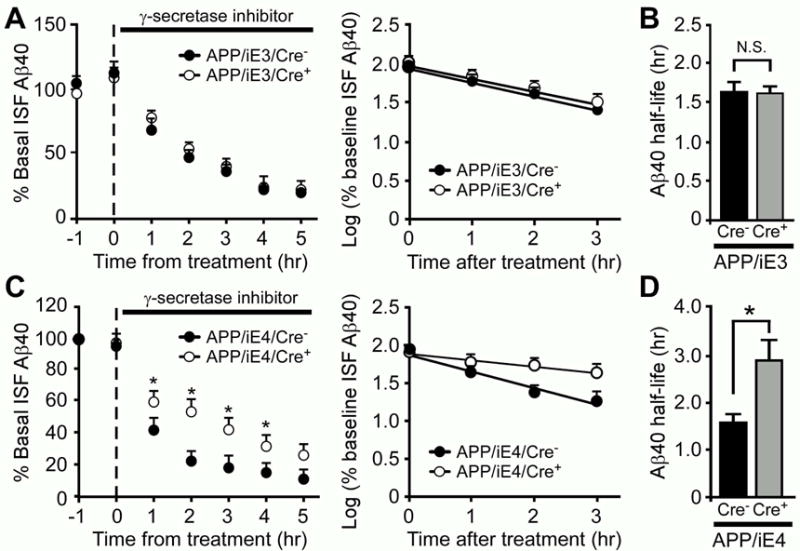
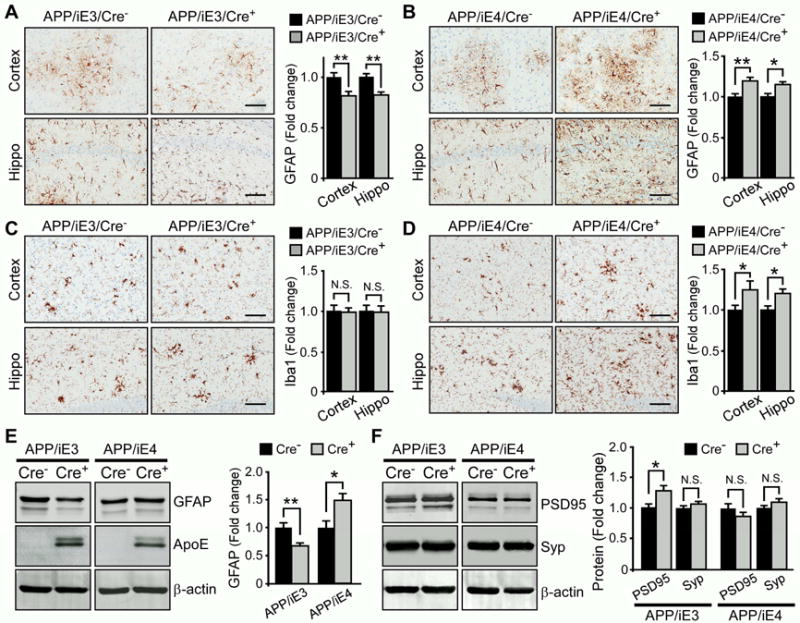
Accumulation and aggregation of amyloid-β (Aβ) in the brain is an initiating step in the pathogenesis of Alzheimer's disease (AD). The ε4 allele of apolipoprotein E (apoE) gene is the strongest genetic risk factor for late-onset AD. Although there is strong evidence showing that apoE4 enhances amyloid pathology, it is not clear what the critical stage(s) is during amyloid development in which apoE4 has the strongest impact. Using apoE inducible mouse models, we show that increased expression of astrocytic apoE4, but not apoE3, during the seeding stage of amyloid development enhanced amyloid deposition and neuritic dystrophy in amyloid model mice. ApoE4, but not apoE3, significantly increased brain Aβ half-life measured by in vivo microdialysis. Furthermore, apoE4 expression increased whereas apoE3 reduced amyloid-related gliosis in the mouse brains. Together, our results demonstrate that apoE4 has the greatest impact on amyloid during the seeding stage, likely by perturbing Aβ clearance and enhancing Aβ aggregation.
Keywords: Alzheimer’s disease; amyloid pathology; apolipoprotein E; seeding.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Seeds of Destruction: New Mechanistic Insights into the Role of Apolipoprotein E4 in Alzheimer's Disease.Neuron. 2017 Dec 6;96(5):953-955. doi: 10.1016/j.neuron.2017.11.022. Neuron. 2017. PMID: 29216455
References
-
- Bales KR, Verina T, Dodel RC, Du Y, Altstiel L, Bender M, Hyslop P, Johnstone EM, Little SP, Cummins DJ, et al. Lack of apolipoprotein E dramatically reduces amyloid beta-peptide deposition. Nat Genet. 1997;17:263–264. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
