

Loss of LOFSEP Transcription Factor Function Converts Spikelet to Leaf-Like Structures in Rice
- PMID: 29217592
- PMCID: PMC5813523
- DOI: 10.1104/pp.17.00704
Loss of LOFSEP Transcription Factor Function Converts Spikelet to Leaf-Like Structures in Rice
Abstract
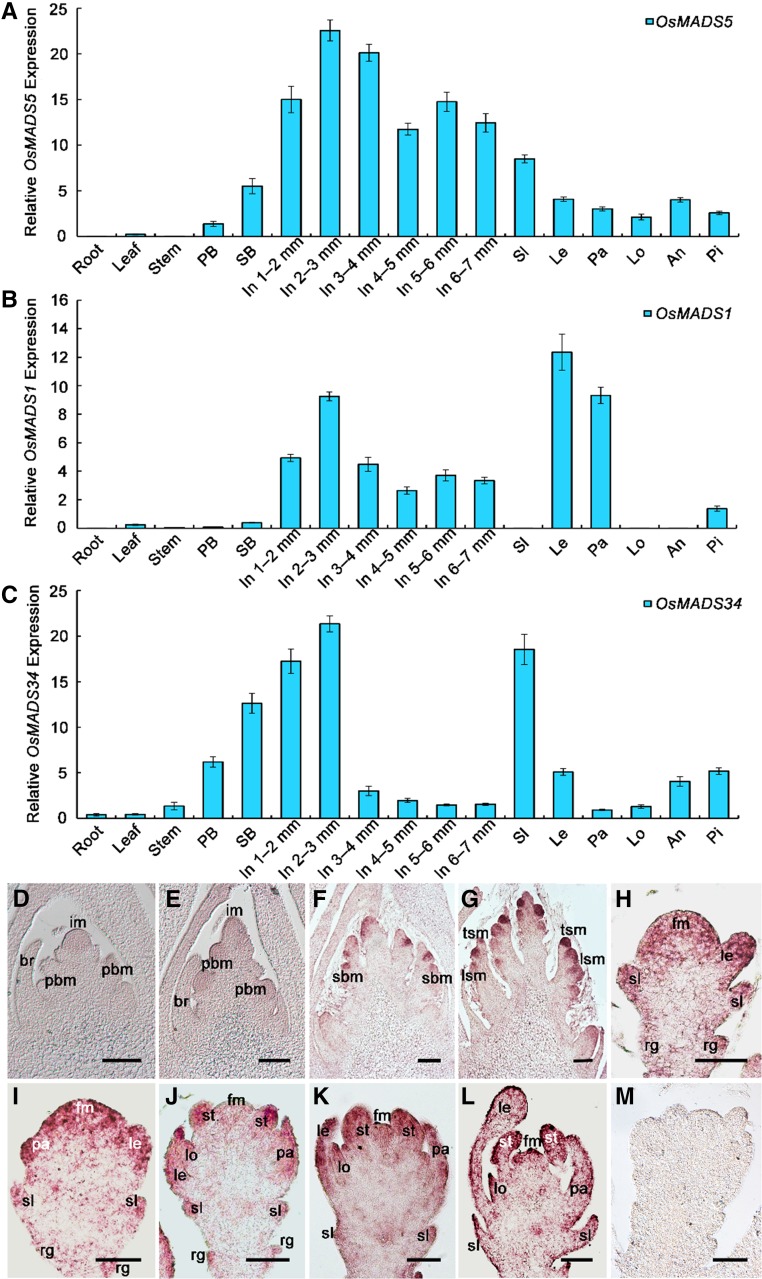
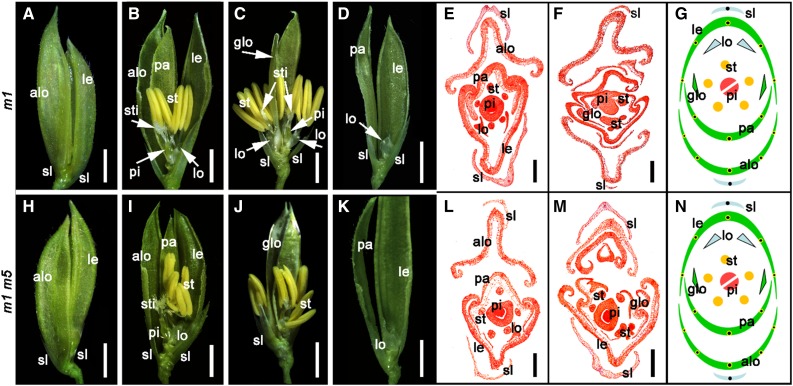
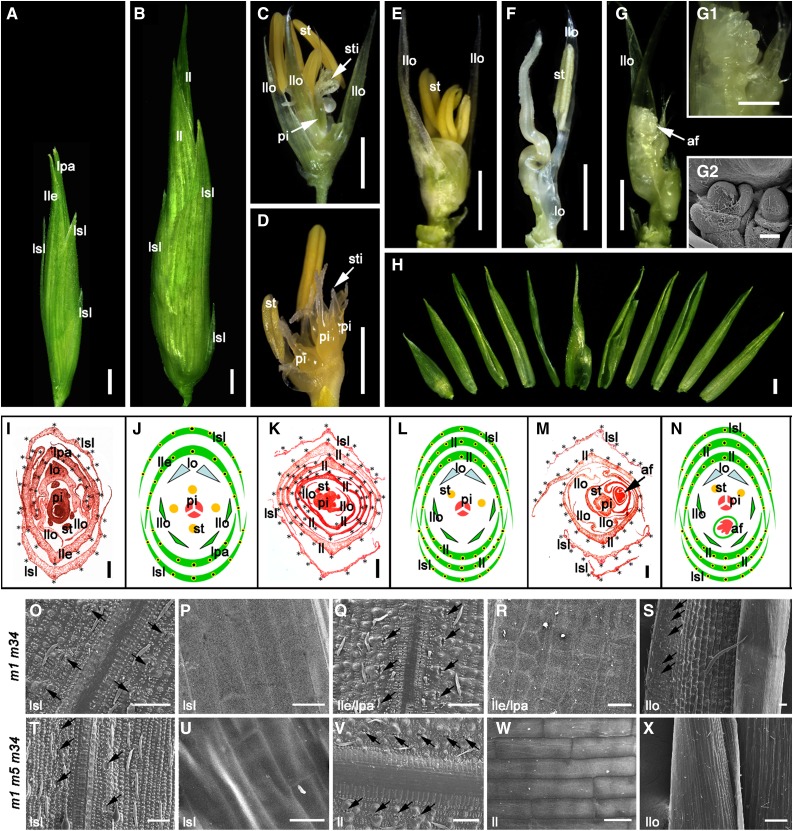
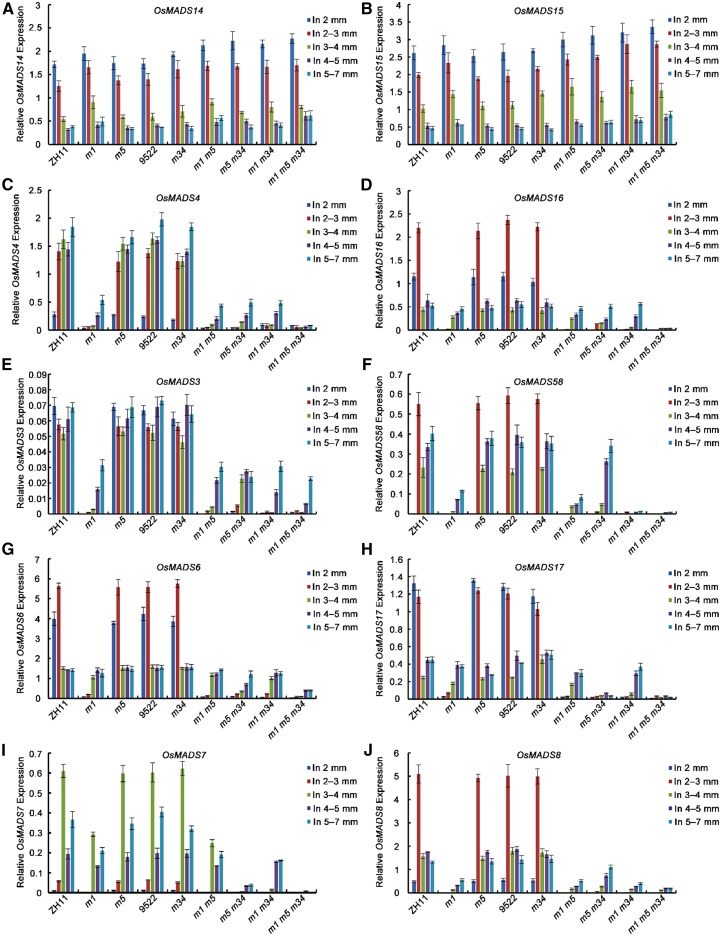
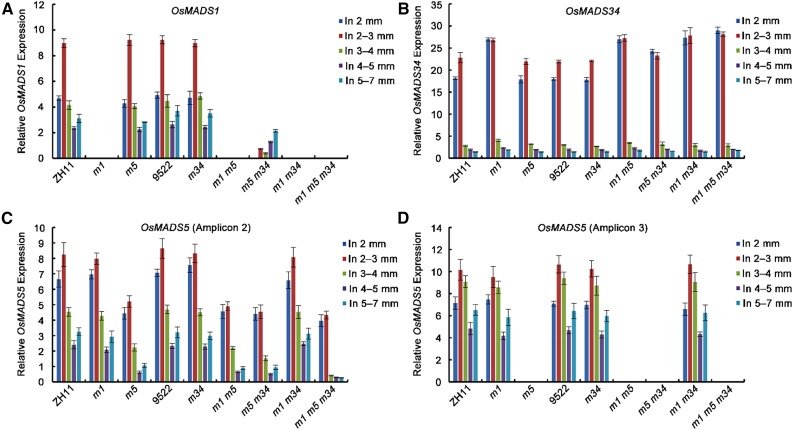
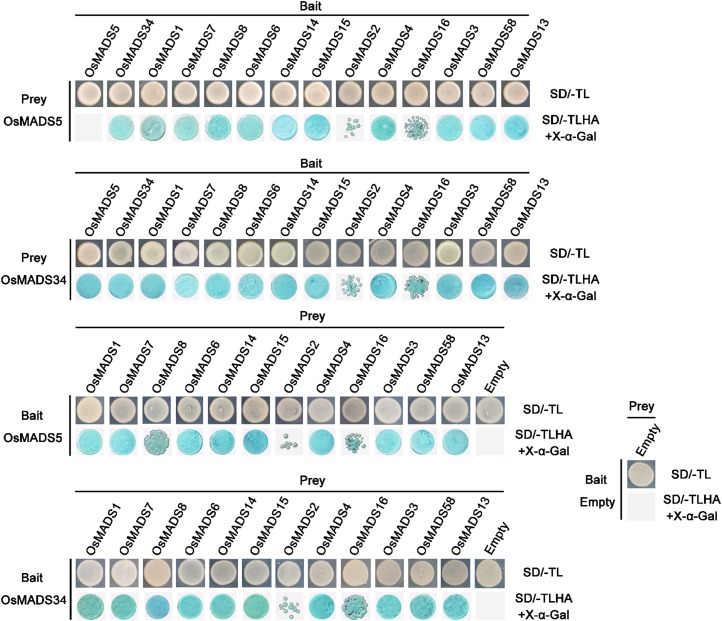
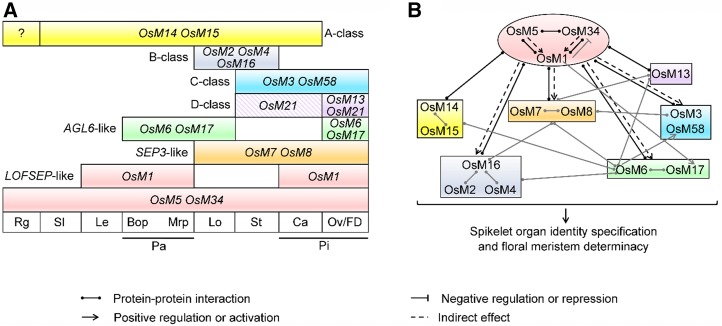
SEPALLATA (SEP)-like genes, which encode a subfamily of MADS-box transcription factors, are essential for specifying floral organ and meristem identity in angiosperms. Rice (Oryza sativa) has five SEP-like genes with partial redundancy and overlapping expression domains, yet their functions and evolutionary conservation are only partially known. Here, we describe the biological role of one of the SEP genes of rice, OsMADS5, in redundantly controlling spikelet morphogenesis. OsMADS5 belongs to the conserved LOFSEP subgroup along with OsMADS1 and OsMADS34OsMADS5 was expressed strongly across a broad range of reproductive stages and tissues. No obvious phenotype was observed in the osmads5 single mutants when compared with the wild type, which was largely due to the functional redundancy among the three LOFSEP genes. Genetic and molecular analyses demonstrated that OsMADS1, OsMADS5, and OsMADS34 together regulate floral meristem determinacy and specify the identities of spikelet organs by positively regulating the other MADS-box floral homeotic genes. Experiments conducted in yeast also suggested that OsMADS1, OsMADS5, and OsMADS34 form protein-protein interactions with other MADS-box floral homeotic members, which seems to be a typical, conserved feature of plant SEP proteins.
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures

References
-
- Agrawal GK, Abe K, Yamazaki M, Miyao A, Hirochika H (2005) Conservation of the E-function for floral organ identity in rice revealed by the analysis of tissue culture-induced loss-of-function mutants of the OsMADS1 gene. Plant Mol Biol 59: 125–135 - PubMed
-
- Causier B, Schwarz-Sommer Z, Davies B (2010) Floral organ identity: 20 years of ABCs. Semin Cell Dev Biol 21: 73–79 - PubMed
-
- Chen ZX, Wu JG, Ding WN, Chen HM, Wu P, Shi CH (2006) Morphogenesis and molecular basis on naked seed rice, a novel homeotic mutation of OsMADS1 regulating transcript level of AP3 homologue in rice. Planta 223: 882–890 - PubMed
-
- Chung YY, Kim SR, Finkel D, Yanofsky MF, An G (1994) Early flowering and reduced apical dominance result from ectopic expression of a rice MADS box gene. Plant Mol Biol 26: 657–665 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
