

Activity-dependent control of NMDA receptor subunit composition at hippocampal mossy fibre synapses
- PMID: 29218821
- PMCID: PMC5813612
- DOI: 10.1113/JP275226
Activity-dependent control of NMDA receptor subunit composition at hippocampal mossy fibre synapses
Abstract
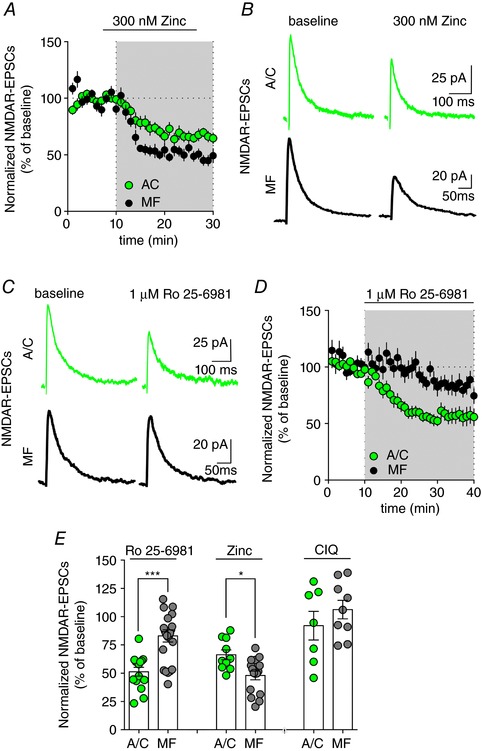
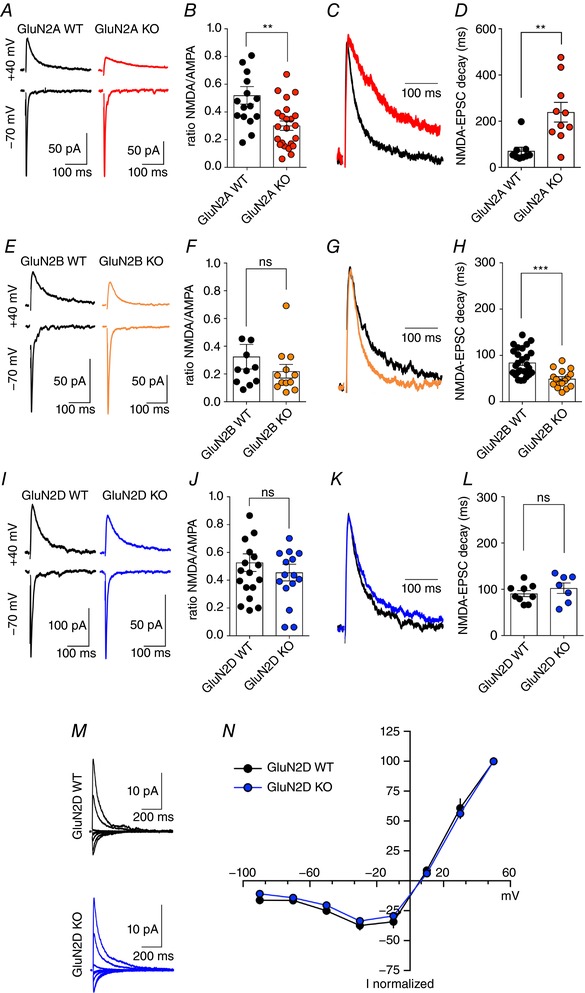
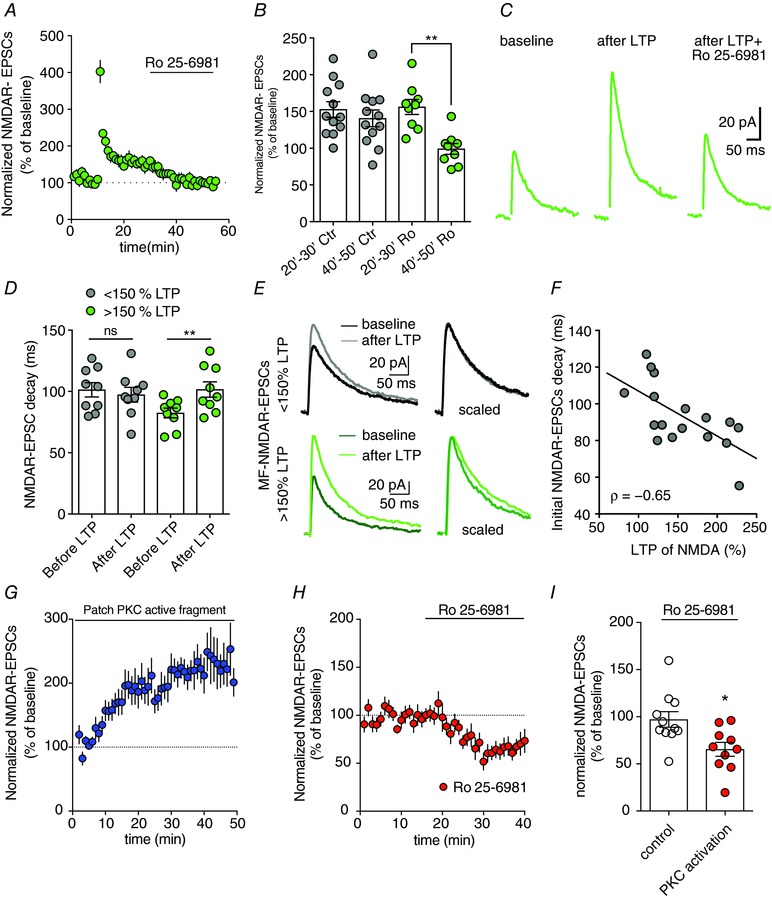
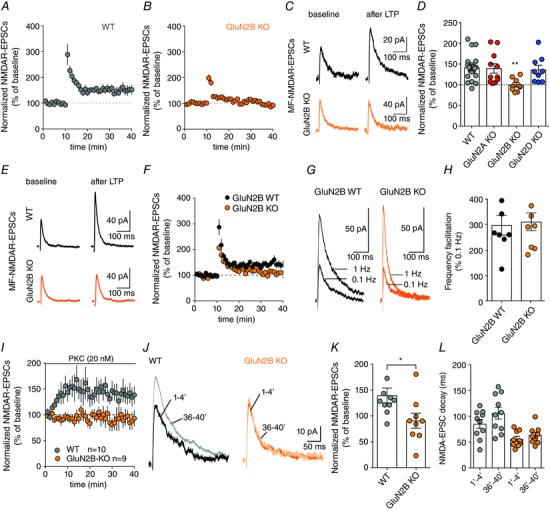
Key points: CA3 pyramidal cells display input-specific differences in the subunit composition of synaptic NMDA receptors (NMDARs). Although at low density, GluN2B contributes significantly to NMDAR-mediated EPSCs at mossy fibre synapses. Long-term potentiation (LTP) of NMDARs triggers a modification in the subunit composition of synaptic NMDARs by insertion of GluN2B. GluN2B subunits are essential for the expression of LTP of NMDARs at mossy fibre synapses.
Abstract: Single neurons express NMDA receptors (NMDARs) with distinct subunit composition and biophysical properties that can be segregated in an input-specific manner. The dynamic control of the heterogeneous distribution of synaptic NMDARs is crucial to control input-dependent synaptic integration and plasticity. In hippocampal CA3 pyramidal cells from mice of both sexes, we found that mossy fibre (MF) synapses display a markedly lower proportion of GluN2B-containing NMDARs than associative/commissural synapses. The mechanism involved in such heterogeneous distribution of GluN2B subunits is not known. Here we show that long-term potentiation (LTP) of NMDARs, which is selectively expressed at MF-CA3 pyramidal cell synapses, triggers a modification in the subunit composition of synaptic NMDARs by insertion of GluN2B. This activity-dependent recruitment of GluN2B at mature MF-CA3 pyramidal cell synapses contrasts with the removal of GluN2B subunits at other glutamatergic synapses during development and in response to activity. Furthermore, although expressed at low levels, GluN2B is necessary for the expression of LTP of NMDARs at MF-CA3 pyramidal cell synapses. Altogether, we reveal a previously unknown activity-dependent regulation and function of GluN2B subunits that may contribute to the heterogeneous plasticity induction rules in CA3 pyramidal cells.
Keywords: NMDA receptors; hippocampus; subunit composition; synaptic plasticity.
© 2017 Centre Nationnal de la Recherche Scientifique. The Journal of Physiology © 2017 The Physiological Society.
Figures
References
-
- Akashi K, Kakizaki T, Kamiya H, Fukaya M, Yamasaki M, Abe M, Natsume R, Watanabe M & Sakimura K (2009). NMDA receptor GluN2B (GluR 2/NR2B) subunit is crucial for channel function, postsynaptic macromolecular organization, and actin cytoskeleton at hippocampal CA3 synapses. J Neurosci 29, 10869–10882. - PMC - PubMed
-
- Barria A & Malinow R (2005). NMDA receptor subunit composition controls synaptic plasticity by regulating binding to CaMKII. Neuron 48, 289–301. - PubMed
-
- Bellone C & Nicoll RA (2007). Rapid bidirectional switching of synaptic NMDA receptors. Neuron 55, 779–785. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous